

Hemorrhagic shock augments lung endothelial cell activation: role of temporal alterations of TLR4 and TLR2
- PMID: 19828841
- PMCID: PMC2803625
- DOI: 10.1152/ajpregu.00445.2009
Hemorrhagic shock augments lung endothelial cell activation: role of temporal alterations of TLR4 and TLR2
Abstract
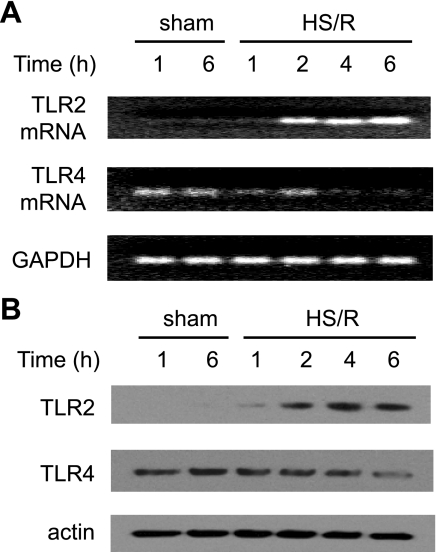
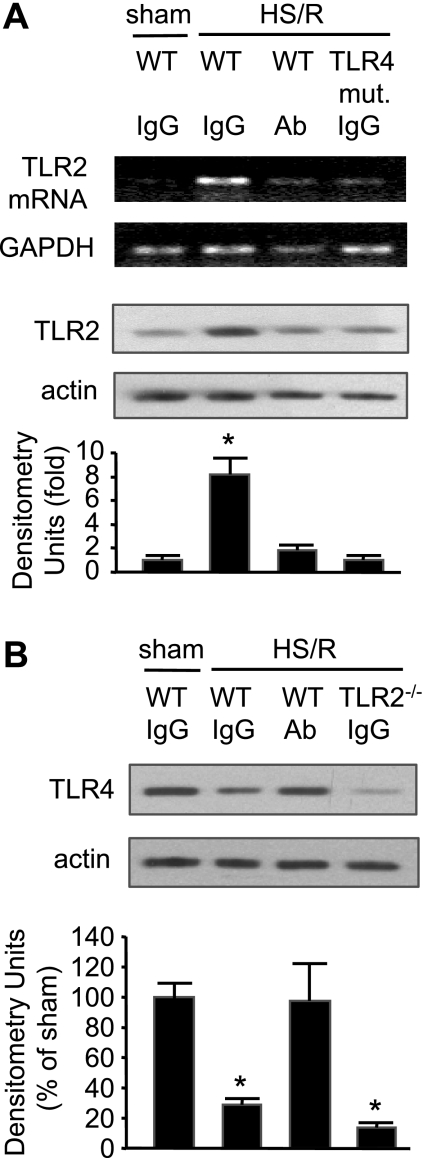
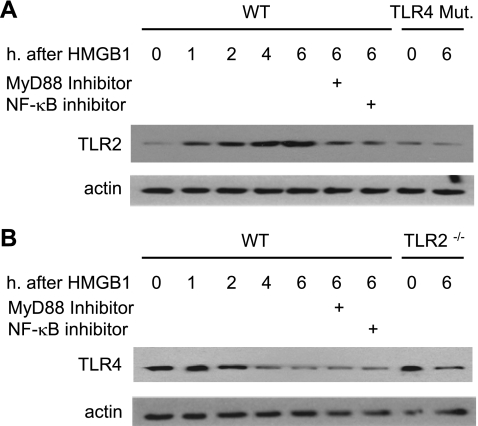
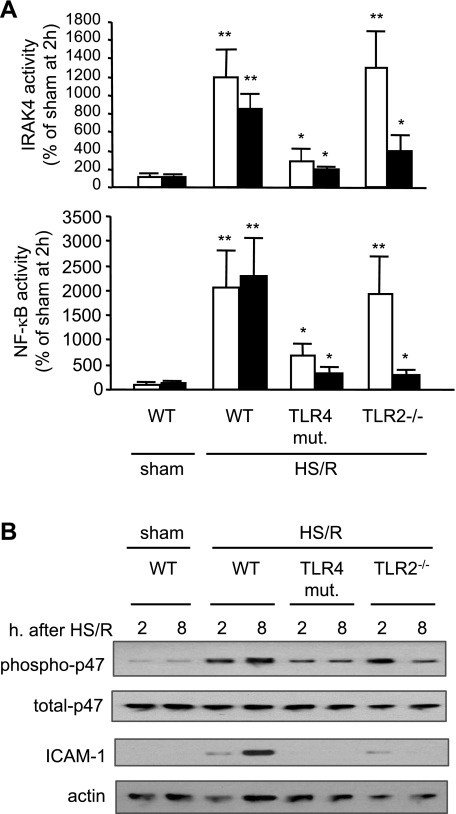
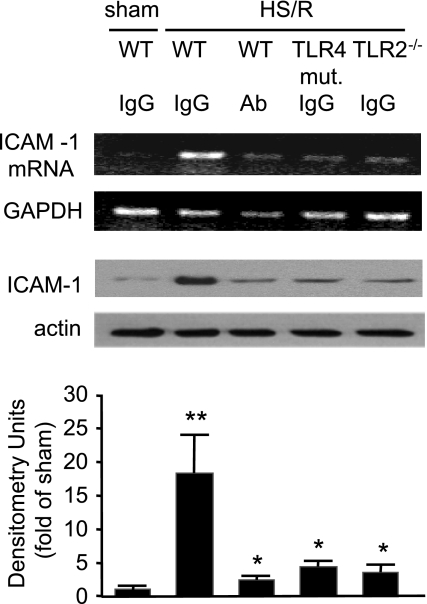
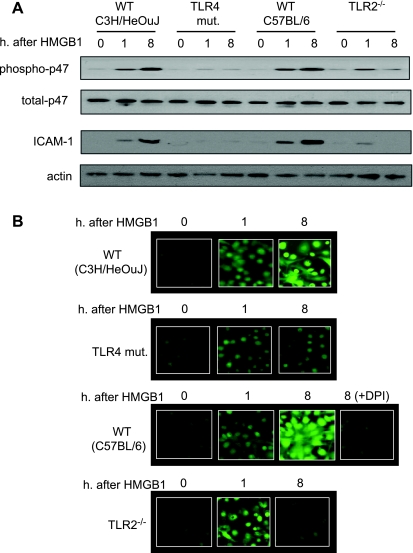
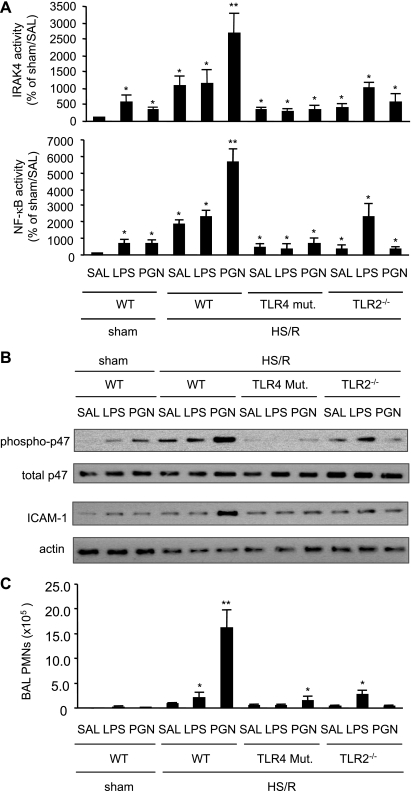
Hemorrhagic shock (HS) due to major trauma predisposes the host to the development of acute lung inflammation and injury. The lung vascular endothelium is an active organ that plays a central role in the development of acute lung injury through generating reactive oxygen species and synthesizing and releasing of a number of inflammatory mediators, including leukocyte adhesion molecules that regulate neutrophils emigration. Previous study from our laboratory has demonstrated that in a setting of sepsis, toll-like receptor-4 (TLR4) signaling can induce TLR2 expression in endothelial cells (ECs), thereby increasing the cells' response to TLR2 ligands. The present study tested the hypothesis that TLR4 activation by HS and the resultant increased TLR2 surface expression in ECs might contribute to the mechanism underlying HS-augmented activation of lung ECs. The results show that high-mobility group box 1 (HMGB1) through TLR4 signaling mediates HS-induced surface expression of TLR2 in the lung and mouse lung vascular endothelial cells (MLVECs). Furthermore, the results demonstrate that HMGB1 induces activation of NAD(P)H oxidase and expression of ICAM-1 in the lung, and MLVECs sequentially depend on TLR4 in the early phase and on TLR2 in the late phase following HS. Finally, the data indicate an important role of the increased TLR2 surface expression in enhancing the activation of MLVECs and augmenting pulmonary neutrophil infiltration in response to TLR2 agonist peptidoglycan. Thus, induction of TLR2 surface expression in lung ECs, induced by HS and mediated by HMGB1/TLR4 signaling, is an important mechanism responsible for endothelial cell-mediated inflammation and organ injury following trauma and hemorrhage.
Figures
References
-
- Akgur FM, Brown MF, Zibari GB, McDonald JC, Epstein CJ, Ross CR, Granger DN. Role of superoxide in hemorrhagic shock-induced P-selectin expression. Am J Physiol Heart Circ Physiol 279: H791– H797, 2000 - PubMed
-
- Akira S, Takeda K. Toll-like receptor signalling. Nat Rev Immunol 4: 499– 511, 2004 - PubMed
-
- Babior BM. The NADPH oxidase of endothelial cells. IUBMB Life 50: 267– 269, 2000 - PubMed
-
- Bonello S, Zahringer C, BelAiba RS, Djordjevic T, Hess J, Michiels C, Kietzmann T, Gorlach A. Reactive oxygen species activate the HIF-1α promoter via a functional NFκB site. Arterioscler Thromb Vasc Biol 27: 755– 761, 2007 - PubMed
-
- Bowie A, O'Neill LA. The interleukin-1 receptor/Toll-like receptor superfamily: signal generators for pro-inflammatory interleukins and microbial products. J Leukoc Biol 67: 508– 514, 2000 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous