

Asymmetric centrosome inheritance maintains neural progenitors in the neocortex
- PMID: 19829375
- PMCID: PMC2764320
- DOI: 10.1038/nature08435
Asymmetric centrosome inheritance maintains neural progenitors in the neocortex
Abstract
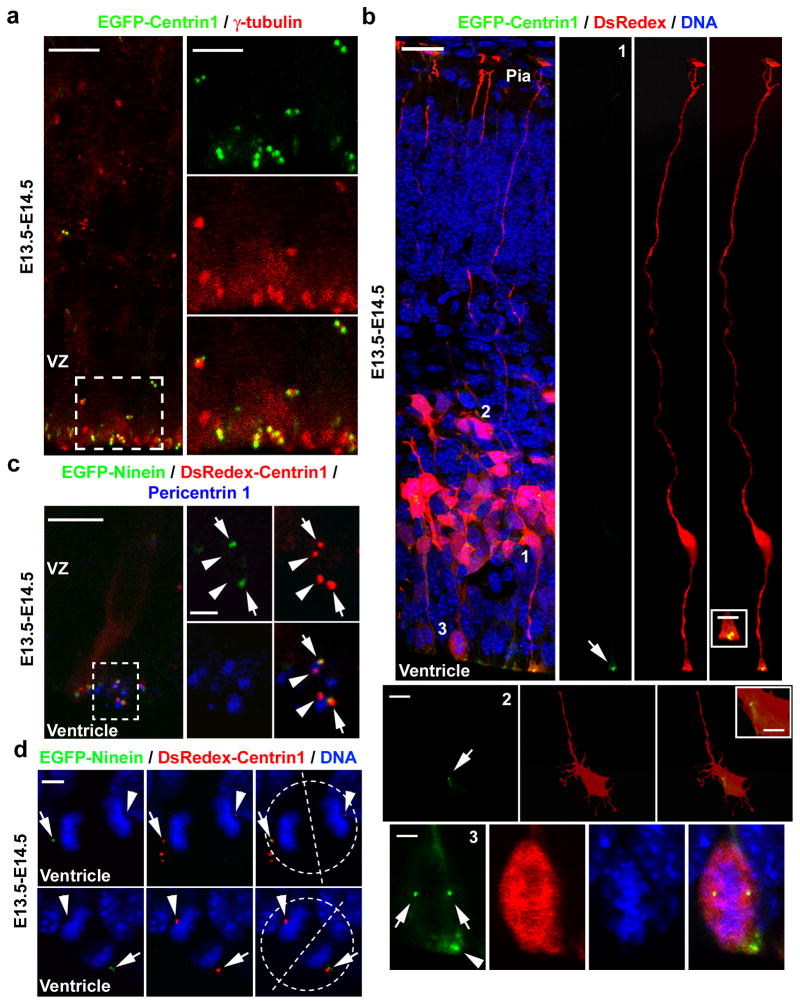
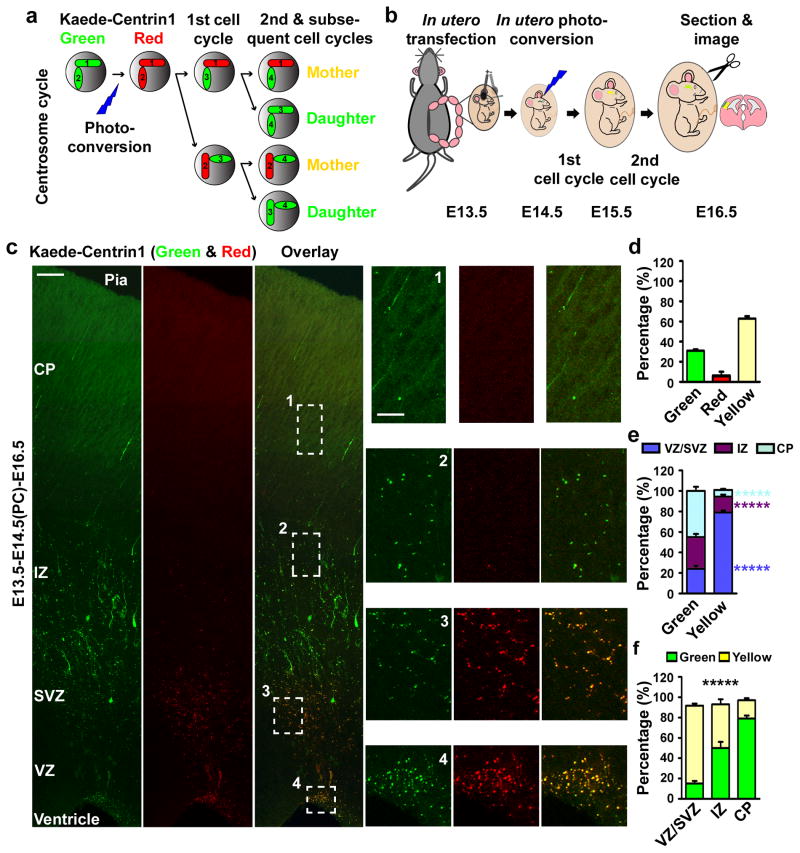
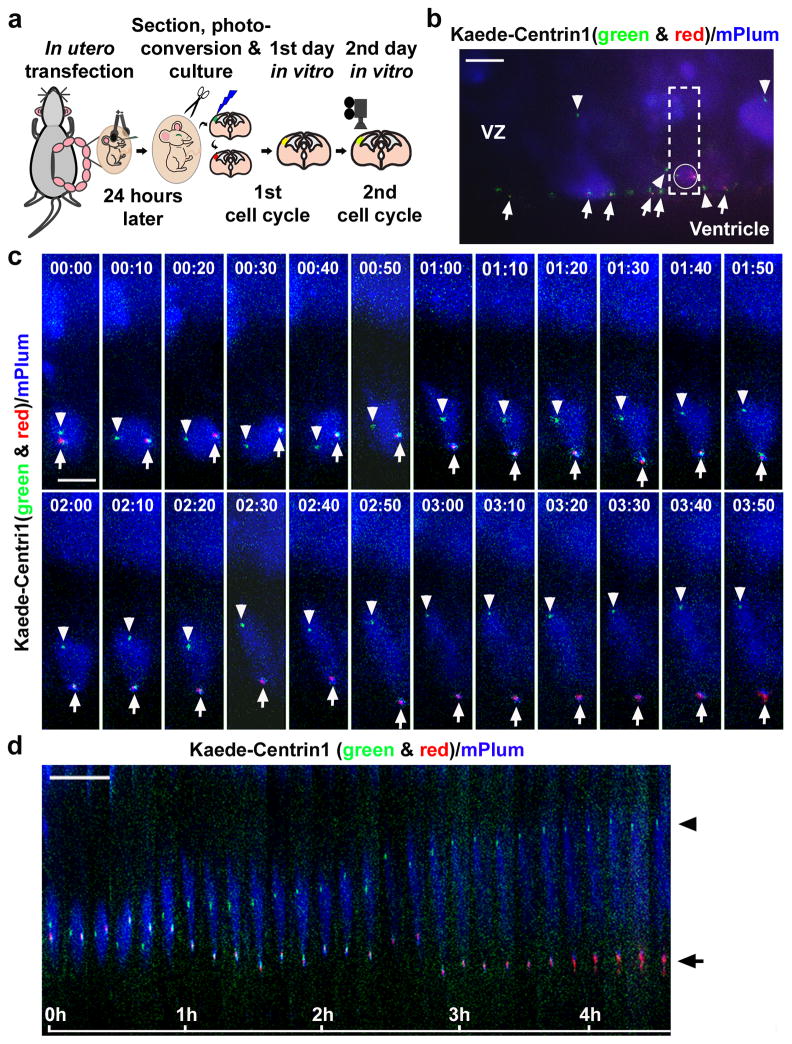
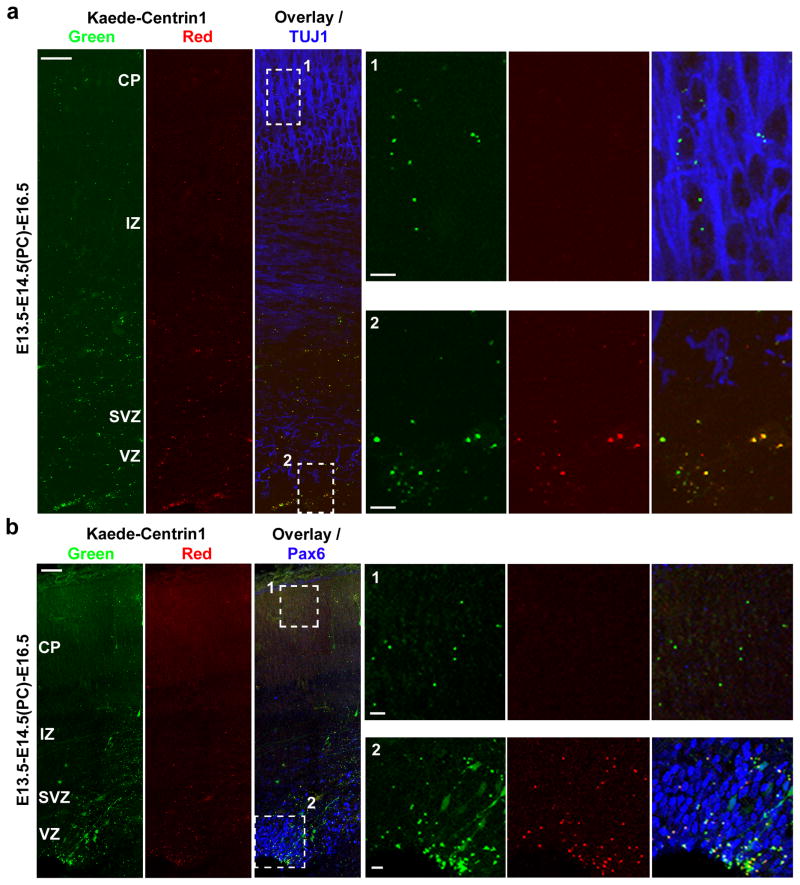
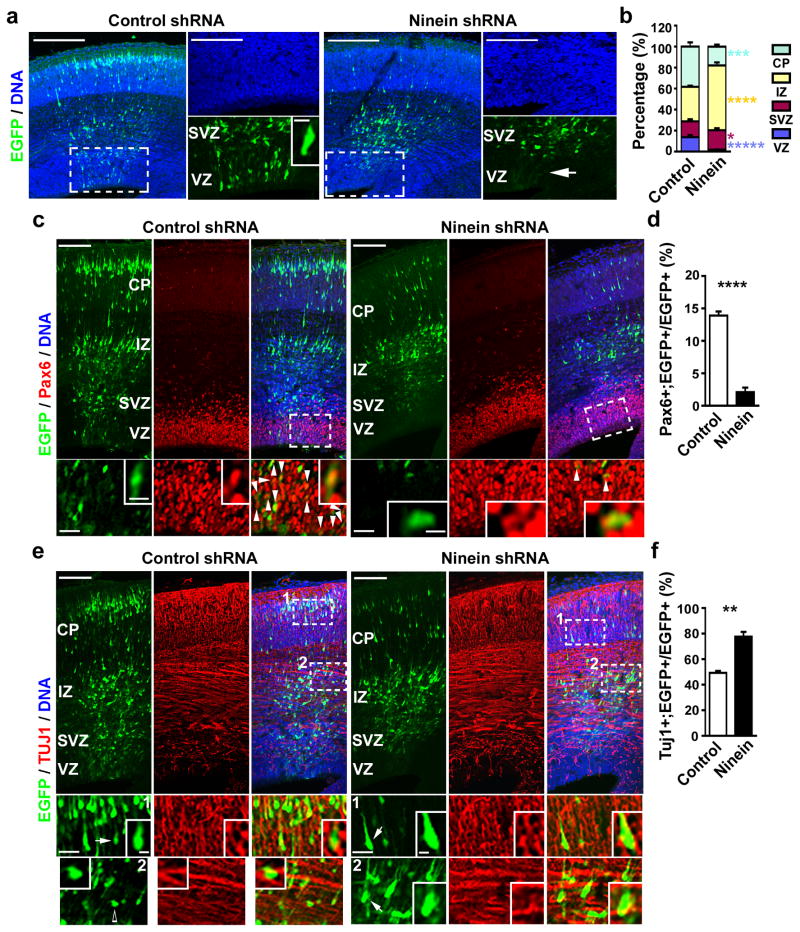
Asymmetric divisions of radial glia progenitors produce self-renewing radial glia and differentiating cells simultaneously in the ventricular zone (VZ) of the developing neocortex. Whereas differentiating cells leave the VZ to constitute the future neocortex, renewing radial glia progenitors stay in the VZ for subsequent divisions. The differential behaviour of progenitors and their differentiating progeny is essential for neocortical development; however, the mechanisms that ensure these behavioural differences are unclear. Here we show that asymmetric centrosome inheritance regulates the differential behaviour of renewing progenitors and their differentiating progeny in the embryonic mouse neocortex. Centrosome duplication in dividing radial glia progenitors generates a pair of centrosomes with differently aged mother centrioles. During peak phases of neurogenesis, the centrosome retaining the old mother centriole stays in the VZ and is preferentially inherited by radial glia progenitors, whereas the centrosome containing the new mother centriole mostly leaves the VZ and is largely associated with differentiating cells. Removal of ninein, a mature centriole-specific protein, disrupts the asymmetric segregation and inheritance of the centrosome and causes premature depletion of progenitors from the VZ. These results indicate that preferential inheritance of the centrosome with the mature older mother centriole is required for maintaining radial glia progenitors in the developing mammalian neocortex.
Figures
Comment in
-
Stem cells: A fateful age gap.Nature. 2009 Oct 15;461(7266):891-2. doi: 10.1038/461891a. Nature. 2009. PMID: 19829363 Free PMC article.
References
-
- Miyata T, Kawaguchi A, Okano H, Ogawa M. Asymmetric inheritance of radial glial fibers by cortical neurons. Neuron. 2001;31:727–41. - PubMed
-
- Noctor SC, Flint AC, Weissman TA, Dammerman RS, Kriegstein AR. Neurons derived from radial glial cells establish radial units in neocortex. Nature. 2001;409:714–20. - PubMed
-
- Malatesta P, Hartfuss E, Gotz M. Isolation of radial glial cells by fluorescent-activated cell sorting reveals a neuronal lineage. Development. 2000;127:5253–63. - PubMed
-
- Rakic P. Elusive radial glial cells: historical and evolutionary perspective. Glia. 2003;43:19–32. - PubMed
-
- Miyata T, et al. Asymmetric production of surface-dividing and non-surface-dividing cortical progenitor cells. Development. 2004;131:3133–45. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
