

Parallel metatranscriptome analyses of host and symbiont gene expression in the gut of the termite Reticulitermes flavipes
- PMID: 19832970
- PMCID: PMC2768689
- DOI: 10.1186/1754-6834-2-25
Parallel metatranscriptome analyses of host and symbiont gene expression in the gut of the termite Reticulitermes flavipes
Abstract
Background: Termite lignocellulose digestion is achieved through a collaboration of host plus prokaryotic and eukaryotic symbionts. In the present work, we took a combined host and symbiont metatranscriptomic approach for investigating the digestive contributions of host and symbiont in the lower termite Reticulitermes flavipes. Our approach consisted of parallel high-throughput sequencing from (i) a host gut cDNA library and (ii) a hindgut symbiont cDNA library. Subsequently, we undertook functional analyses of newly identified phenoloxidases with potential importance as pretreatment enzymes in industrial lignocellulose processing.
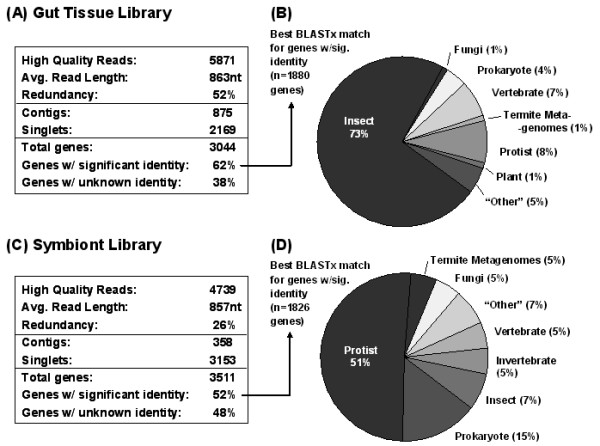
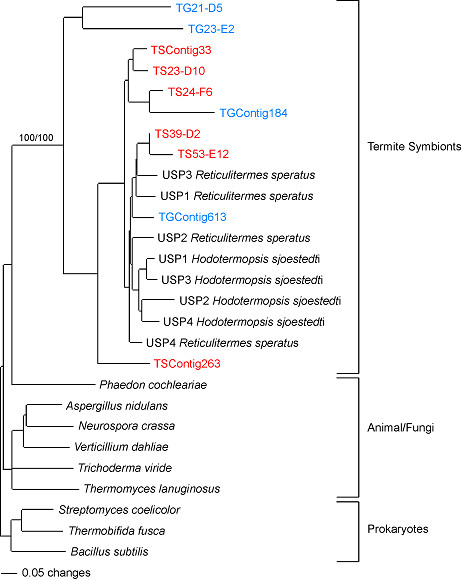
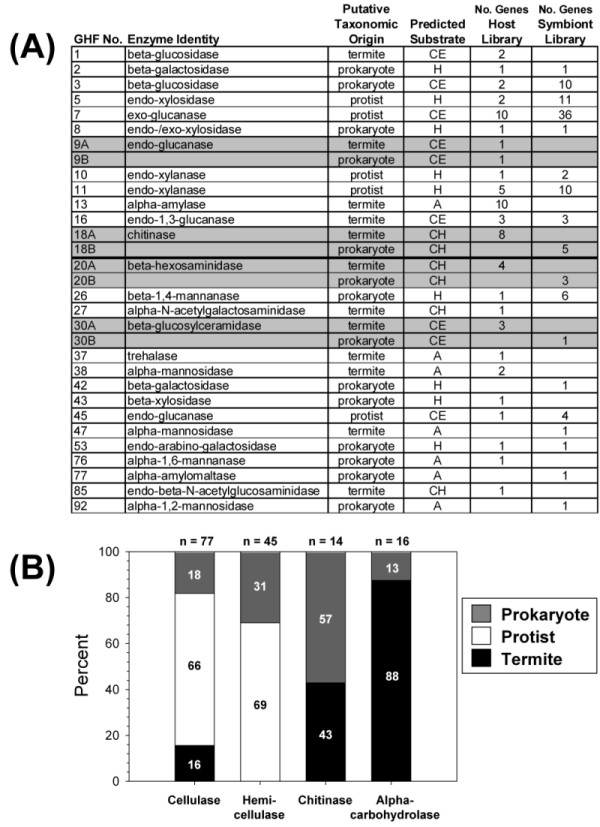
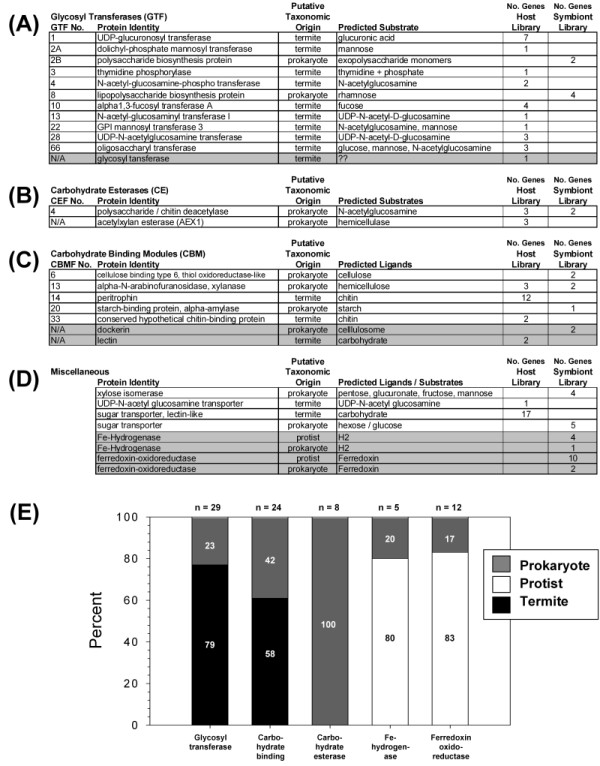
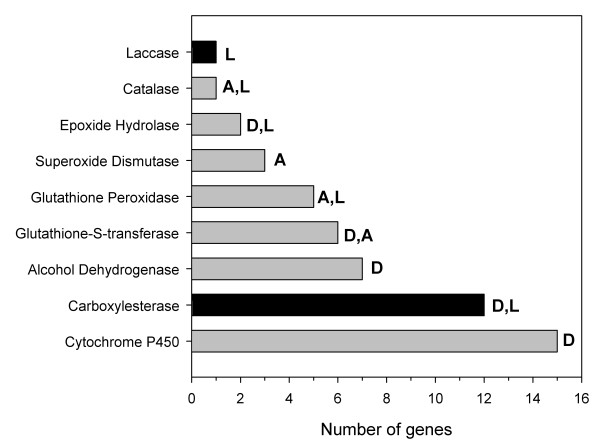
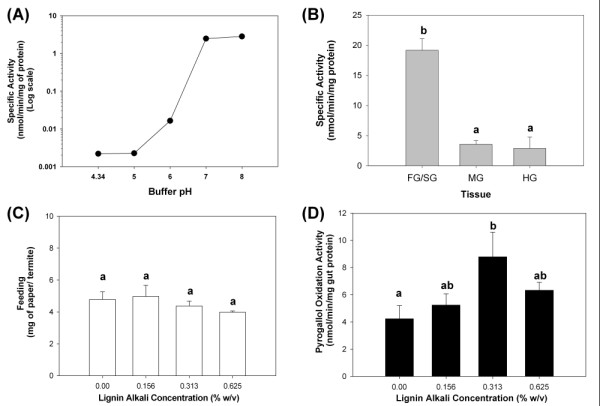
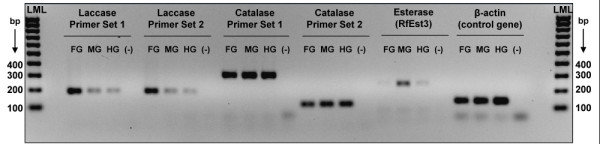
Results: Over 10,000 expressed sequence tags (ESTs) were sequenced from the 2 libraries that aligned into 6,555 putative transcripts, including 171 putative lignocellulase genes. Sequence analyses provided insights in two areas. First, a non-overlapping complement of host and symbiont (prokaryotic plus protist) glycohydrolase gene families known to participate in cellulose, hemicellulose, alpha carbohydrate, and chitin degradation were identified. Of these, cellulases are contributed by host plus symbiont genomes, whereas hemicellulases are contributed exclusively by symbiont genomes. Second, a diverse complement of previously unknown genes that encode proteins with homology to lignase, antioxidant, and detoxification enzymes were identified exclusively from the host library (laccase, catalase, peroxidase, superoxide dismutase, carboxylesterase, cytochrome P450). Subsequently, functional analyses of phenoloxidase activity provided results that were strongly consistent with patterns of laccase gene expression. In particular, phenoloxidase activity and laccase gene expression are mostly restricted to symbiont-free foregut plus salivary gland tissues, and phenoloxidase activity is inducible by lignin feeding.
Conclusion: To our knowledge, this is the first time that a dual host-symbiont transcriptome sequencing effort has been conducted in a single termite species. This sequence database represents an important new genomic resource for use in further studies of collaborative host-symbiont termite digestion, as well as development of coevolved host and symbiont-derived biocatalysts for use in industrial biomass-to-bioethanol applications. Additionally, this study demonstrates that: (i) phenoloxidase activities are prominent in the R. flavipes gut and are not symbiont derived, (ii) expands the known number of host and symbiont glycosyl hydrolase families in Reticulitermes, and (iii) supports previous models of lignin degradation and host-symbiont collaboration in cellulose/hemicellulose digestion in the termite gut. All sequences in this paper are available publicly with the accession numbers FL634956-FL640828 (Termite Gut library) and FL641015-FL645753 (Symbiont library).
Figures
References
-
- Scharf ME, Tartar A. Termite digestomes as sources for novel lignocellulases. Biofuels Bioprod Bioref. 2008;2:540–552. doi: 10.1002/bbb.107. - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
