

Transcriptome analysis of functional differentiation between haploid and diploid cells of Emiliania huxleyi, a globally significant photosynthetic calcifying cell
- PMID: 19832986
- PMCID: PMC2784329
- DOI: 10.1186/gb-2009-10-10-r114
Transcriptome analysis of functional differentiation between haploid and diploid cells of Emiliania huxleyi, a globally significant photosynthetic calcifying cell
Abstract
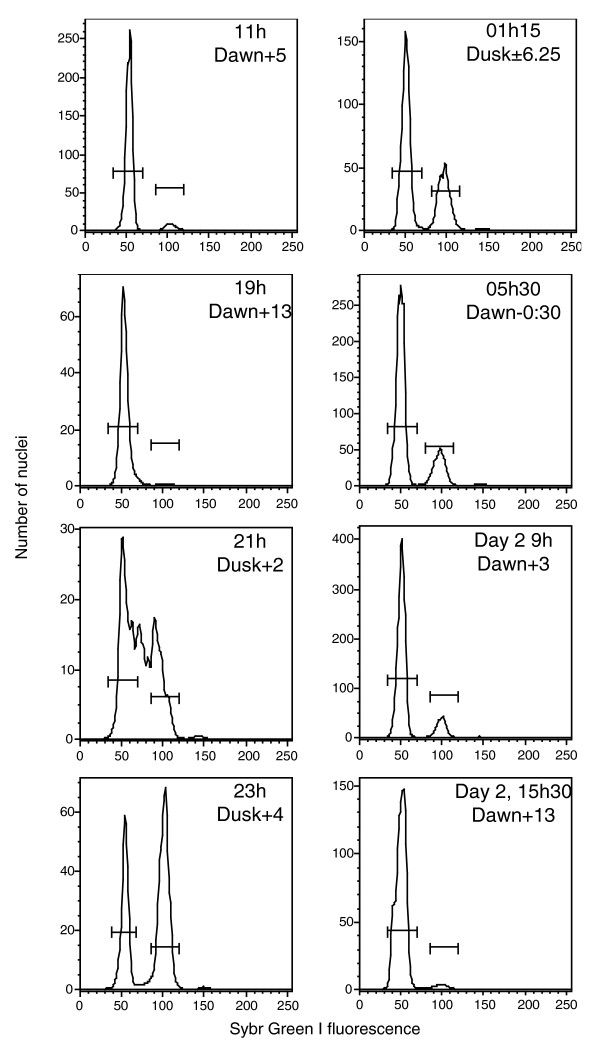
Background: Eukaryotes are classified as either haplontic, diplontic, or haplo-diplontic, depending on which ploidy levels undergo mitotic cell division in the life cycle. Emiliania huxleyi is one of the most abundant phytoplankton species in the ocean, playing an important role in global carbon fluxes, and represents haptophytes, an enigmatic group of unicellular organisms that diverged early in eukaryotic evolution. This species is haplo-diplontic. Little is known about the haploid cells, but they have been hypothesized to allow persistence of the species between the yearly blooms of diploid cells. We sequenced over 38,000 expressed sequence tags from haploid and diploid E. huxleyi normalized cDNA libraries to identify genes involved in important processes specific to each life phase (2N calcification or 1N motility), and to better understand the haploid phase of this prominent haplo-diplontic organism.
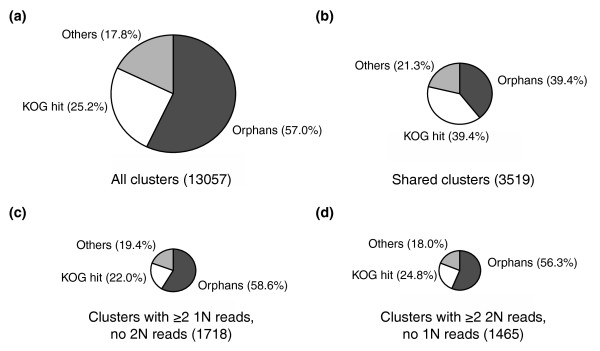
Results: The haploid and diploid transcriptomes showed a dramatic differentiation, with approximately 20% greater transcriptome richness in diploid cells than in haploid cells and only <or= 50% of transcripts estimated to be common between the two phases. The major functional category of transcripts differentiating haploids included signal transduction and motility genes. Diploid-specific transcripts included Ca2+, H+, and HCO3- pumps. Potential factors differentiating the transcriptomes included haploid-specific Myb transcription factor homologs and an unusual diploid-specific histone H4 homolog.
Conclusions: This study permitted the identification of genes likely involved in diploid-specific biomineralization, haploid-specific motility, and transcriptional control. Greater transcriptome richness in diploid cells suggests they may be more versatile for exploiting a diversity of rich environments whereas haploid cells are intrinsically more streamlined.
Figures








References
-
- de Vargas C, Aubry M-P, Probert I, Young J. In: Evolution of aquatic photoautotrophs. Falkowski PG, Knoll AH, editor. New York: Academic Press; 2007. Origin and evolution of coccolithophores: From coastal hunters to oceanic farmers. pp. 251–285.
-
- Andersen RA. Biology and systematics of heterokont and haptophyte algae. Am J Bot. 2004;91:1508–1522. - PubMed
-
- Young JR, Geisen M, Probert I. A review of selected aspects of coccolithophore biology with implications for paleodiversity estimation. Micropaleontology. 2005;51:1–22.
-
- Sanchez-Puerta MV, Delwiche CF. A hypothesis for plastid evolution in chromalveolates. J Phycol. 2008;44:1097–1107. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
