

Kinetics of early T cell receptor signaling regulate the pathway of lytic granule delivery to the secretory domain
- PMID: 19833088
- PMCID: PMC2778196
- DOI: 10.1016/j.immuni.2009.09.004
Kinetics of early T cell receptor signaling regulate the pathway of lytic granule delivery to the secretory domain
Abstract
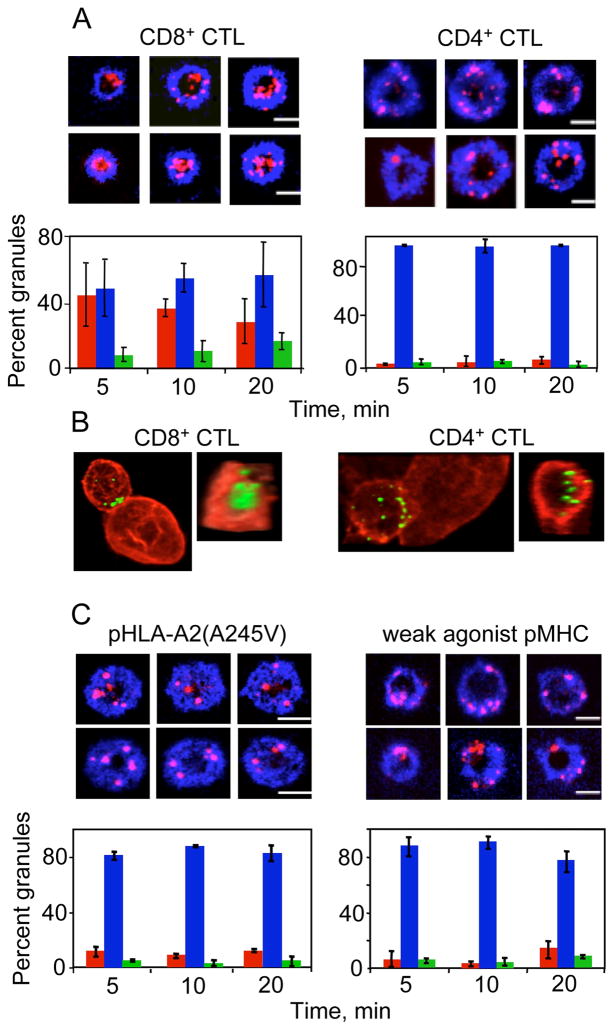
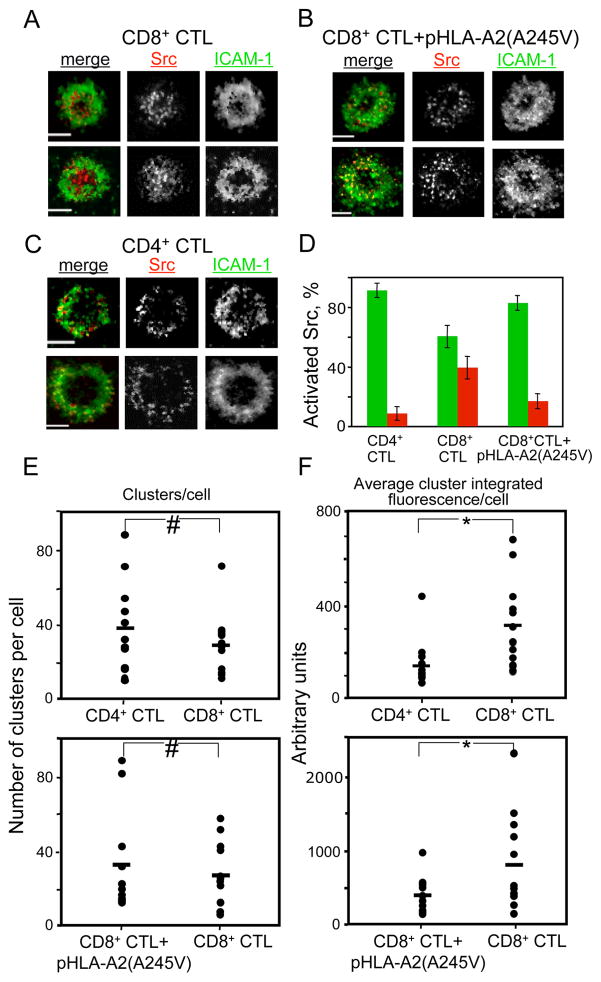
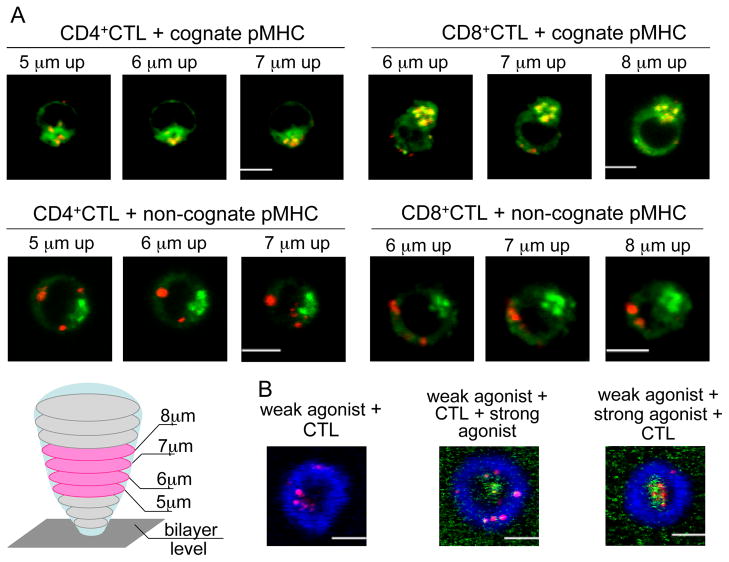
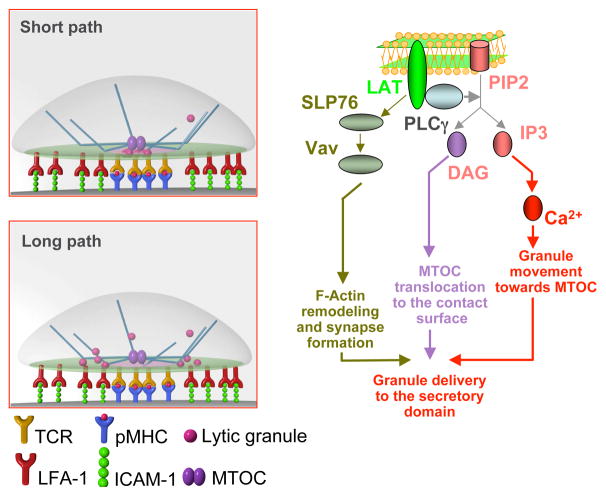
Cytolytic granules mediate killing of virus-infected cells by cytotoxic T lymphocytes. We show here that the granules can take long or short paths to the secretory domain. Both paths utilized the same intracellular molecular events, which have different spatial and temporal arrangements and are regulated by the kinetics of Ca(2+)-mediated signaling. Rapid signaling caused swift granule concentration near the microtubule-organizing center (MTOC) and subsequent delivery by the polarized MTOC directly to the secretory domain-the shortest path. Indolent signaling led to late recruitment of granules that moved along microtubules to the periphery of the synapse and then moved tangentially to fuse at the outer edge of the secretory domain-a longer path. The short pathway is associated with faster granule release and more efficient killing than the long pathway. Thus, the kinetics of early signaling regulates the quality of the T cell cytolytic response.
Figures



Comment in
-
A view to a kill: how ligand quality controls lethal hits.Immunity. 2009 Oct 16;31(4):531-3. doi: 10.1016/j.immuni.2009.09.011. Immunity. 2009. PMID: 19833082
References
-
- Appay V, Zaunders JJ, Papagno L, Sutton J, Jaramillo A, Waters A, Easterbrook P, Grey P, Smith D, McMichael AJ, et al. Characterization of CD4(+) CTLs ex vivo. J Immunol. 2002;168:5954–5958. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
