

Review
doi: 10.1098/rstb.2009.0131.
Effects of heat stress on mammalian reproduction
Affiliations
- PMID: 19833646
- PMCID: PMC2781849
- DOI: 10.1098/rstb.2009.0131
Item in Clipboard
Review
Effects of heat stress on mammalian reproduction
Philos Trans R Soc Lond B Biol Sci.
.
Abstract
Heat stress can have large effects on most aspects of reproductive function in mammals. These include disruptions in spermatogenesis and oocyte development, oocyte maturation, early embryonic development, foetal and placental growth and lactation. These deleterious effects of heat stress are the result of either the hyperthermia associated with heat stress or the physiological adjustments made by the heat-stressed animal to regulate body temperature. Many effects of elevated temperature on gametes and the early embryo involve increased production of reactive oxygen species. Genetic adaptation to heat stress is possible both with respect to regulation of body temperature and cellular resistance to elevated temperature.
Figures
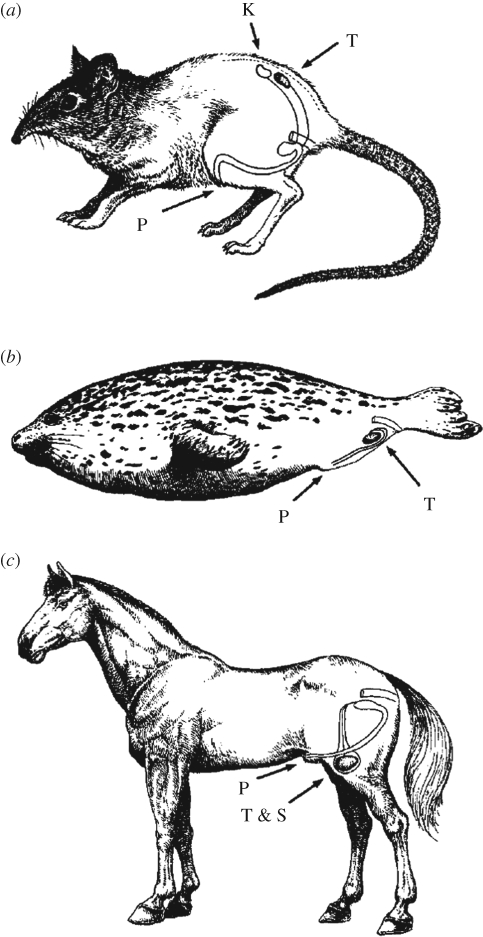
Variation in the location of the testes in mammals. Most mammals have testes that are located outside the abdominal cavity and that exist at a temperature a few degrees cooler than core body temperature. One example is the (c) horse. However, in some species, descent of the testis from a location caudal to the kidneys does not occur or is very limited and testes remain in the dorsal abdominal cavity. The (a) rofous elephant shrew, Elephantulus rufescens, is an example. For other species, the testes descend to the ventral abdominal wall but remain in the body cavity, as is the case for the (b) harbour seal, Phoca vitulina. T, testis; K, kidney; P, penis; T & S, testis in scrotum. The figure is adapted from Werdelin & Nilsonne (1999) with permission from Journal of Theoretical Biology.
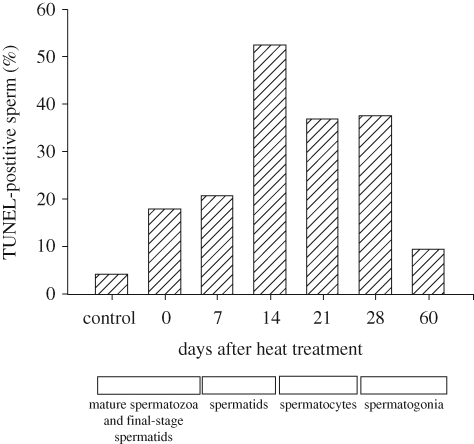
DNA damage in epididymal sperm collected at various times after heating of the scrotum of male mice. DNA damage was determined using the TUNEL assay. The x-axis indicates the days after heating when the sample was collected and the approximate stage at which recovered spermatogenic cells were when heating was applied. The figure is redrawn from Pérez-Crespo et al. (2008) with permission from Molecular Reproduction and Development.
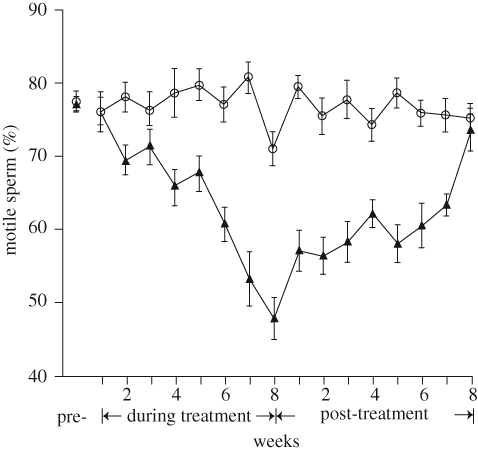
Time course of effects of heat stress on motility of ejaculated spermatozoa in bulls. Control bulls (circles) were maintained in a chamber at 23 ± 1°C for 16 weeks. Heat-stressed bulls (triangles) were maintained for 8 weeks in a chamber that was at 31 ± 1°C for 8 h and 35 ± 1°C for 16 h of each 24 h day. Thereafter, heat-stressed bulls were returned to an environment of 23 ± 1°C for 8 weeks. The figure is adapted from Meyerhoeffer et al. (1985) with permission from Journal of Animal Science.
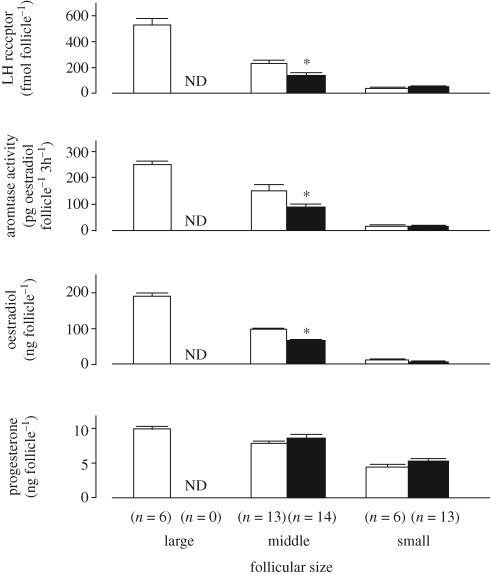
Effects of heat stress on characteristics of follicles during the peri-ovulatory period in goats. Goats were heat stressed (black bar) or not (open bar) for 48 h, treated with prostaglandin F-2α and after 48 additional hours, follicles that had been recruited in the past 24 h were collected and analysed. ND, not determined; *, p < 0.05. The figure is adapted from Ozawa et al. (2005) with permission from Reproduction.
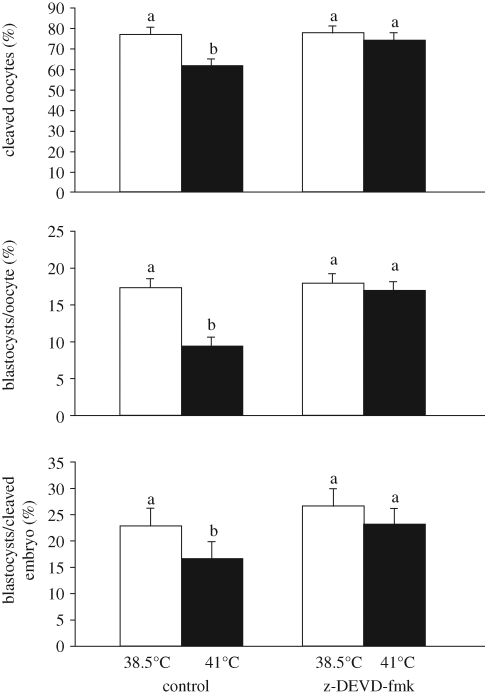
Inhibition of apoptosis with the caspase inhibitor z-DEVD-fmk reduces the effects of heat shock on oocyte maturation. Bovine oocytes were matured at either 38.5°C or 41°C in the presence or absence of z-DEVD-fmk. Subsequently, oocytes were fertilized and the resultant embryos cultured until determination of blastocyst development at day 8 after insemination. Note that the effects of heat shock during maturation on cleavage and the proportion of oocytes or cleaved embryos that became blastocysts was reduced in the presence of the inhibitor. The figure is adapted from Roth & Hansen (2004a) with permission from Biology of Reproduction.
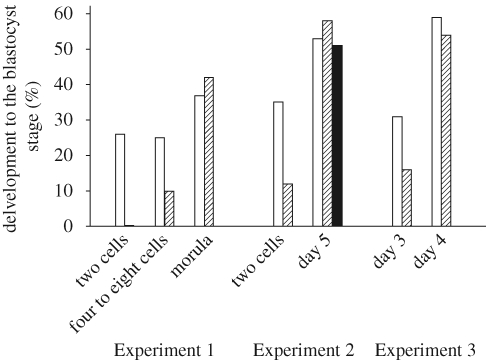
Bovine preimplantation embryos become more resistant to heat shock as development advances. Shown are the results of three experiments (Experiment 1, Edwards & Hansen 1997; Experiment 2, Krininger et al. 2002; Experiment 3, Sakatani et al. 2004) where embryos at various stages of development or days after insemination were cultured at 38.5–39°C (open bars) or at an elevated culture temperature of 41°C for 12 h (hatched bar, Experiment 1), 41°C for 6 h (hatched bar) or 9 h (filled bar) (Experiment 2) or 43°C for 1 h (hatched, Experiment 3). The figure is adapted from Hansen (2007) with permission from Theriogenology.
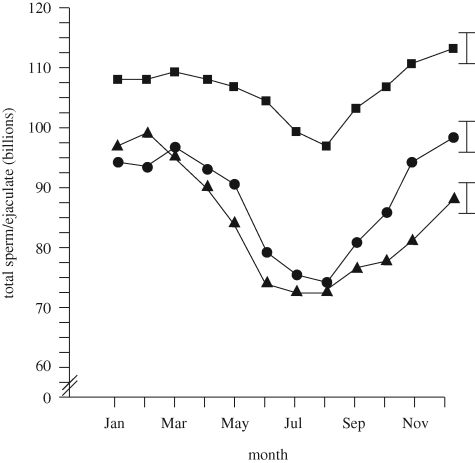
Differences between genetic lines of boars in seasonal variation in sperm output per ejaculate. Square, genetic line A; triangle, genetic line B; circle, genetic line C. The figure is adapted from Flowers (2008) with permission from Theriogenology.
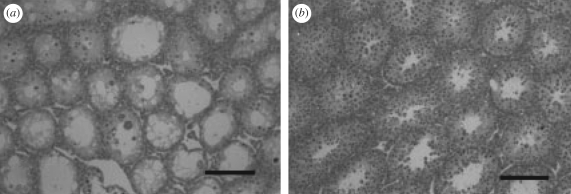
Strain differences in mice for testicular changes following surgical cryptorchidism. Shown are cross sections of testicular tissue from (a) C57/BL6 and (b) MRL/Mpj mice. Note that the seminiferous tubules were larger in MRL/mpj mice and that germ cells were more abundant including spermatocytes and spermatids. The figure is adapte from Kon & Endoh (2001) with permission from Reproduction, Fertility and Development.
References
-
- Alexander G., Hales J. R. S., Stevens D., Donnelly J. B.1987Effects of acute and prolonged exposure to heat on regional blood flows in pregnant sheep. J. Dev. Physiol. 9, 1–15 - PubMed
-
- Al-Katanani Y. M., Webb D. W., Hansen P. J.1999Factors affecting seasonal variation in 90 day non-return rate to first service in lactating Holstein cows in a hot climate. J Dairy Sci. 82, 2611–2615 - PubMed
-
- Al-Katanani Y. M., Paula-Lopes F. F., Hansen P. J.2002Effect of season and exposure to heat stress on oocyte competence in Holstein cows. J. Dairy Sci. 85, 390–396 - PubMed
-
- Aréchiga C. F., Hansen P. J.1998Response of preimplantation murine embryos to heat shock as modified by developmental stage and glutathione status. In Vitro Cell Dev. Biol. Anim. 34, 655–659 (doi:10.1007/s11626-996-0016-8) - DOI - PubMed
-
- Aréchiga C. F., Ealy A. D., Hansen P. J.1995Evidence that glutathione is involved in thermotolerance of preimplantation mouse embryos. Biol. Reprod. 52, 1296–1301 (doi:10.1095/biolreprod52.6.1296) - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources