

Methionine oxidation contributes to bacterial killing by the myeloperoxidase system of neutrophils
- PMID: 19833874
- PMCID: PMC2774013
- DOI: 10.1073/pnas.0909464106
Methionine oxidation contributes to bacterial killing by the myeloperoxidase system of neutrophils
Abstract
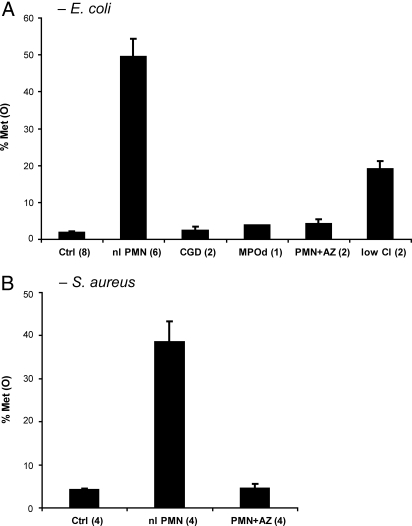
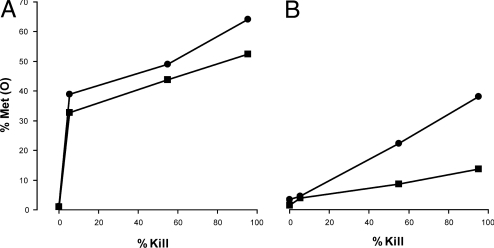
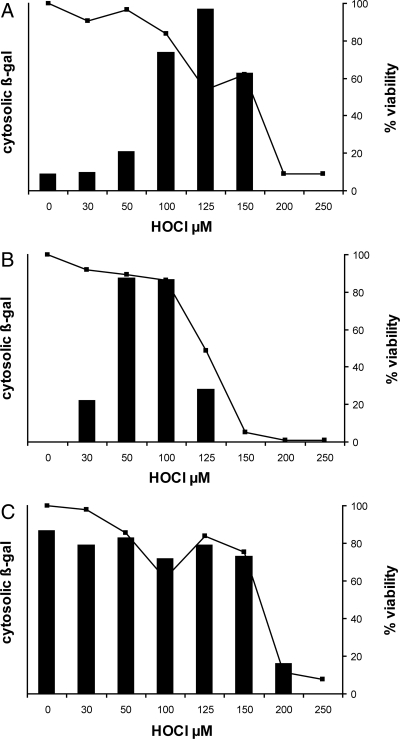
Reactive oxygen intermediates generated by neutrophils kill bacteria and are implicated in inflammatory tissue injury, but precise molecular targets are undefined. We demonstrate that neutrophils use myeloperoxidase (MPO) to convert methionine residues of ingested Escherichia coli to methionine sulfoxide in high yield. Neutrophils deficient in individual components of the MPO system (MPO, H(2)O(2), chloride) exhibited impaired bactericidal activity and impaired capacity to oxidize methionine. HOCl, the principal physiologic product of the MPO system, is a highly efficient oxidant for methionine, and its microbicidal effects were found to correspond linearly with oxidation of methionine residues in bacterial cytosolic and inner membrane proteins. In contrast, outer envelope proteins were initially oxidized without associated microbicidal effect. Disruption of bacterial methionine sulfoxide repair systems rendered E. coli more susceptible to killing by HOCl, whereas over-expression of a repair enzyme, methionine sulfoxide reductase A, rendered them resistant, suggesting a direct role for methionine oxidation in bactericidal activity. Prominent among oxidized bacterial proteins were those engaged in synthesis and translocation of peptides to the cell envelope, an essential physiological function. Moreover, HOCl impaired protein translocation early in the course of bacterial killing. Together, our findings indicate that MPO-mediated methionine oxidation contributes to bacterial killing by neutrophils. The findings further suggest that protein translocation to the cell envelope is one important pathway targeted for damage.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Nauseef WM. How human neutrophils kill and degrade microbes: An integrated view. Immunol Rev. 2007;219:88–102. - PubMed
-
- Klebanoff SJ. Myeloperoxidase: Friend and foe. J Leukoc Biol. 2005;77:598–625. - PubMed
-
- Klebanoff SJ, Clark RA. The Neutrophil. Amsterdam: North-Holland Publishing; 1978.
-
- Harrison JE, Schultz J. Studies on the chlorinating activity of myeloperoxidase. J Biol Chem. 1976;251:1371–1374. - PubMed
-
- Chapman AL, Hampton MB, Senthilmohan R, Winterbourn CC, Kettle AJ. Chlorination of bacterial and neutrophil proteins during phagocytosis and killing of Staphylococcus aureus. J Biol Chem. 2002;277:9757–9762. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
