

Increased synthesis of leukotrienes in the mouse model of diabetic retinopathy
- PMID: 19834040
- PMCID: PMC2868429
- DOI: 10.1167/iovs.09-3557
Increased synthesis of leukotrienes in the mouse model of diabetic retinopathy
Abstract
Purpose: Evidence suggests that capillary degeneration in early diabetic retinopathy results from chronic inflammation, and leukotrienes have been implicated in this process. The authors investigated the cellular sources of leukotriene biosynthesis in diabetic retinas and the effects of hyperglycemia on leukotriene production.
Methods: Retinas and bone marrow cells were collected from diabetic and nondiabetic mice. Mouse retinal glial cells and retinal endothelial cells (mRECs) were cultured under nondiabetic and diabetic conditions. Production of leukotriene metabolites was assessed by mass spectrometry, and Western blot analysis was used to quantitate the expression of enzymes and receptors involved in leukotriene synthesis and signaling.
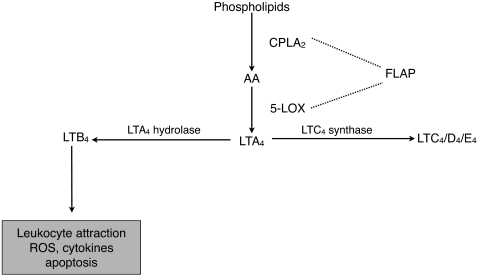
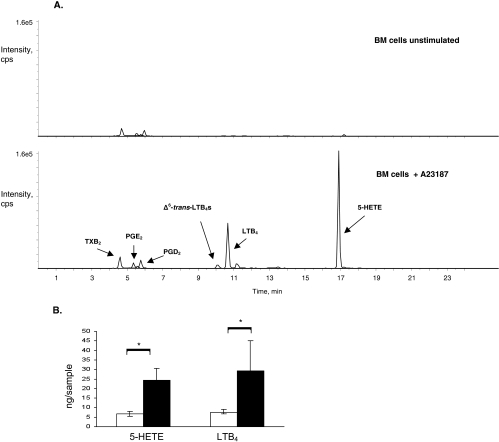
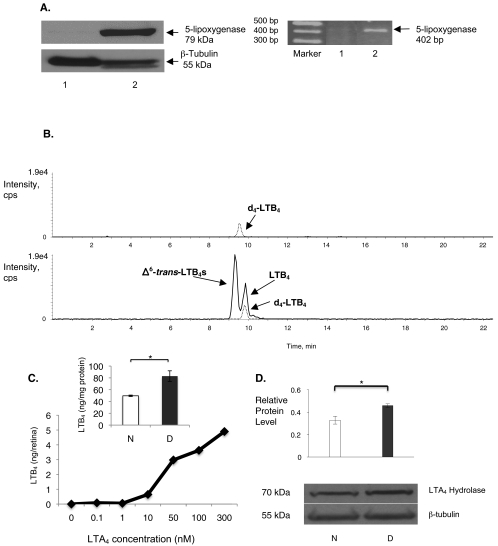
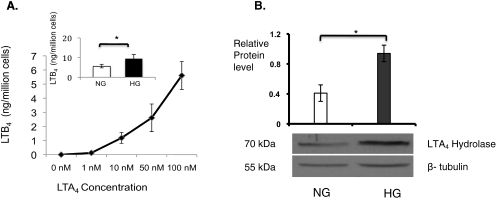
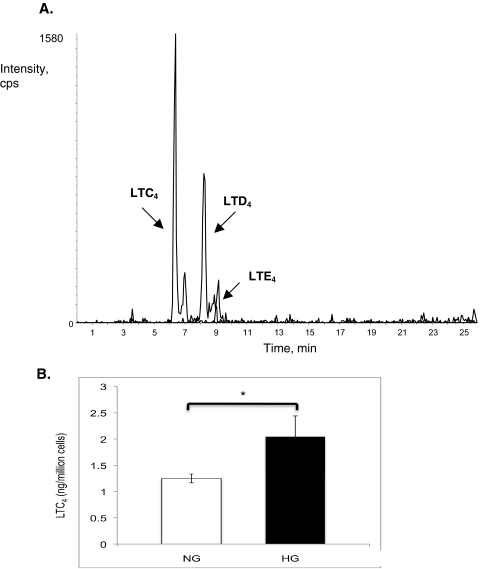
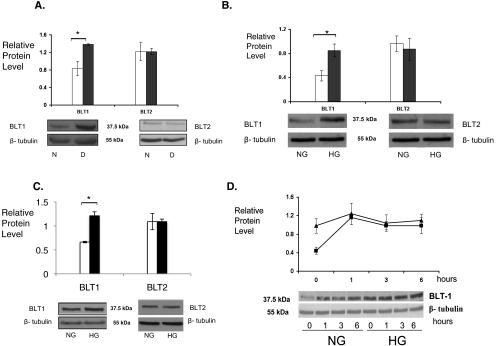
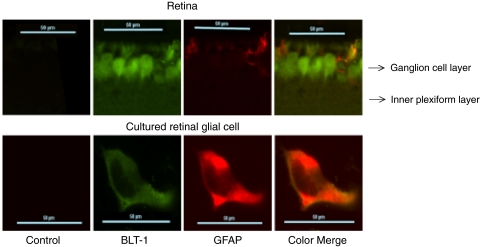
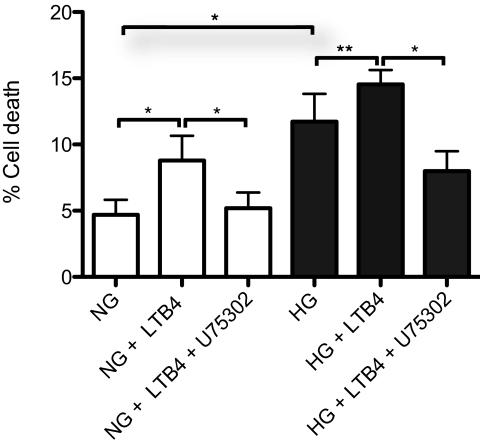
Results: Bone marrow cells from nondiabetic mice expressed 5-lipoxygenase, the enzyme required for the initiation of leukotriene synthesis, and produced leukotriene B(4) (LTB(4)) when stimulated with the calcium ionophore A23187. Notably, LTB(4) synthesis was increased threefold over normal (P < 0.03) in bone marrow cells from diabetic mice. In contrast, retinas from nondiabetic or diabetic mice produced neither leukotrienes nor 5-lipoxygenase mRNA. Despite an inability to initiate leukotriene biosynthesis, the addition of exogenous leukotriene A(4) (LTA(4); the precursor of LTB(4)) to retinas resulted in robust production of LTB(4). Similarly, retinal glial cells synthesized LTB(4) from LTA(4), whereas mRECs produced both LTB(4) and the cysteinyl leukotrienes. Culturing the retinal cells in high-glucose concentrations enhanced leukotriene synthesis and selectively increased expression of the LTB(4) receptor BLT1. Antagonism of the BLT1 receptor inhibited LTB(4)-induced mREC cell death.
Conclusions: Transcellular delivery of LTA(4) from marrow-derived cells to retinal cells results in the generation of LTB(4) and the death of endothelial cells and, thus, might contribute to chronic inflammation and retinopathy in diabetes.
Figures
References
-
- Antonetti DA, Barber AJ, Khin S, Lieth E, Tarbell JM, Gardner TW. Vascular permeability in experimental diabetes is associated with reduced endothelial occludin content: vascular endothelial growth factor decreases occludin in retinal endothelial cells. Penn State Retina Research Group Diabetes 1998;47:1953–1959 - PubMed
-
- Antonetti DA, Lieth E, Barber AJ, Gardner TW. Molecular mechanisms of vascular permeability in diabetic retinopathy. Semin Ophthalmol 1999;14:240–248 - PubMed
-
- Du Y, Miller CM, Kern TS. Hyperglycemia increases mitochondrial superoxide in retina and retinal cells. Free Radical Biol Med 2003;35:1491–1499 - PubMed
-
- Gardner TW, Antonetti DA, Barber AJ, LaNoue KF, Nakamura M. New insights into the pathophysiology of diabetic retinopathy: potential cell-specific therapeutic targets. Diabetes Technol Ther 2000;2:601–608 - PubMed
-
- Joussen AM, Poulaki V, Le ML, et al. A central role for inflammation in the pathogenesis of diabetic retinopathy. FASEB J 2004;18:1450–1452 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
