

Schlank, a member of the ceramide synthase family controls growth and body fat in Drosophila
- PMID: 19834458
- PMCID: PMC2790492
- DOI: 10.1038/emboj.2009.305
Schlank, a member of the ceramide synthase family controls growth and body fat in Drosophila
Abstract
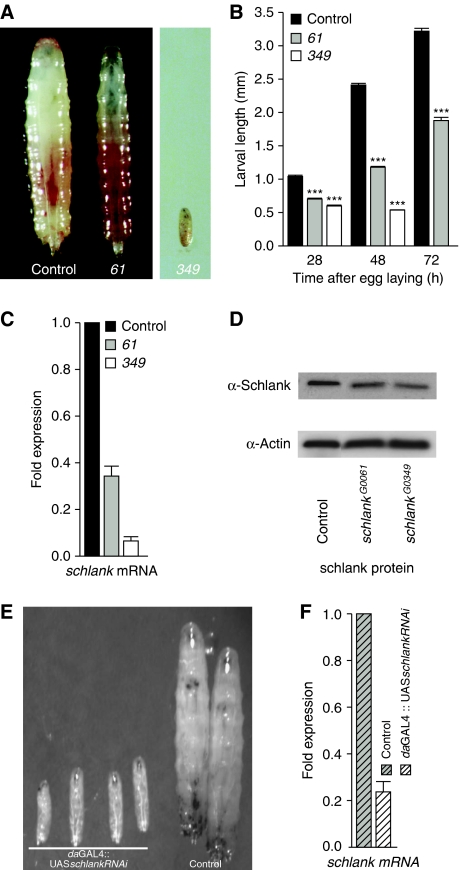
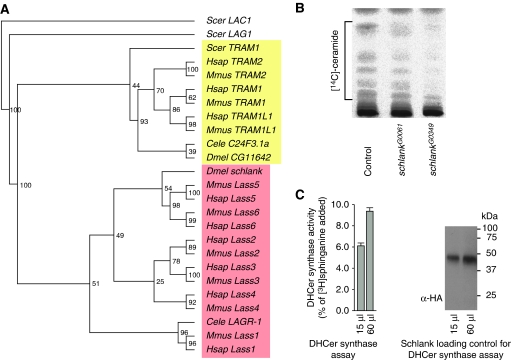
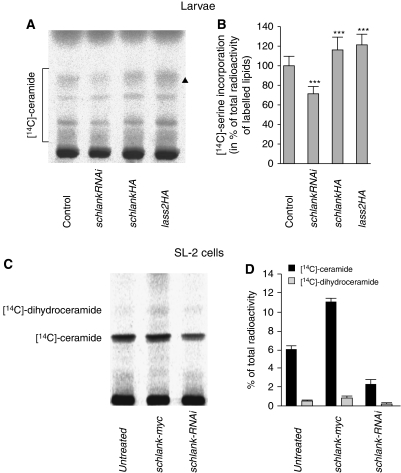
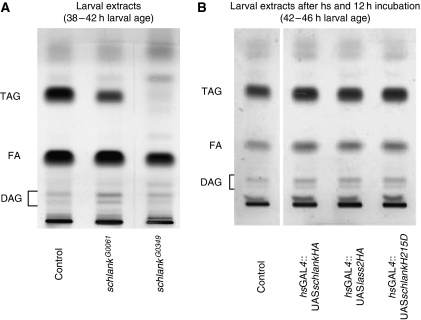
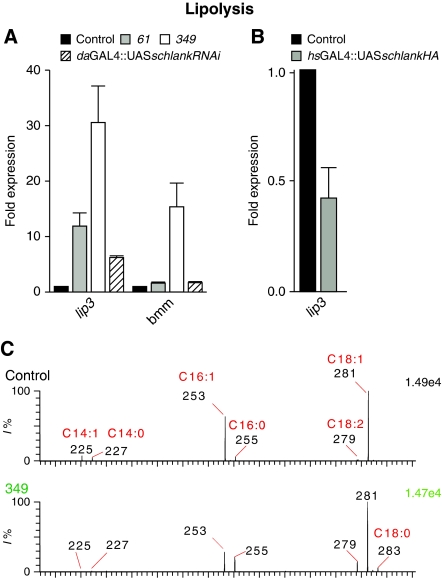
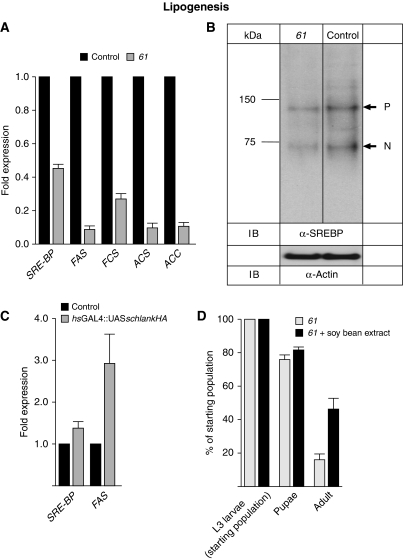
Ceramide synthases are highly conserved transmembrane proteins involved in the biosynthesis of sphingolipids, which are essential structural components of eukaryotic membranes and can act as second messengers regulating tissue homeostasis. However, the role of these enzymes in development is poorly understood due to the lack of animal models. We identified schlank as a new Drosophila member of the ceramide synthase family. We demonstrate that schlank is involved in the de novo synthesis of a broad range of ceramides, the key metabolites of sphingolipid biosynthesis. Unexpectedly, schlank mutants also show reduction of storage fat, which is deposited as triacylglyerols in the fat body. We found that schlank can positively regulate fatty acid synthesis by promoting the expression of sterol-responsive element-binding protein (SREBP) and SREBP-target genes. It further prevents lipolysis by downregulating the expression of triacylglycerol lipase. Our results identify schlank as a new regulator of the balance between lipogenesis and lipolysis in Drosophila. Furthermore, our studies of schlank and the mammalian Lass2 family member suggest a novel role for ceramide synthases in regulating body fat metabolism.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Aguila JR, Suszko J, Gibbs AG, Hoshizaki DK (2007) The role of larval fat cells in adult Drosophila melanogaster. J Exp Biol 210: 956–963 - PubMed
-
- Arrese EL, Wells MA (1997) Adipokinetic hormone induced lipolysis in the fat body of an insect, Manduca sexta: synthesis of sn-1,2-diacylglycerols. J Lipid Res 38: 68–76 - PubMed
-
- Batheja AD, Uhlinger DJ, Carton JM, Ho G, D'Andrea MR (2003) Characterization of serine palmitoyltransferase in normal human tissues. J Histochem Cytochem 51: 687–696 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
