

Induction of membrane ceramides: a novel strategy to interfere with T lymphocyte cytoskeletal reorganisation in viral immunosuppression
- PMID: 19834551
- PMCID: PMC2757718
- DOI: 10.1371/journal.ppat.1000623
Induction of membrane ceramides: a novel strategy to interfere with T lymphocyte cytoskeletal reorganisation in viral immunosuppression
Abstract
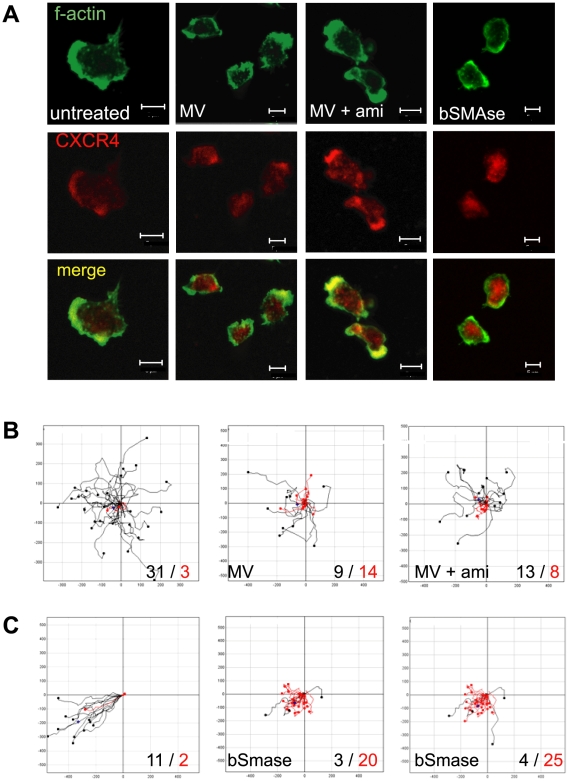
Silencing of T cell activation and function is a highly efficient strategy of immunosuppression induced by pathogens. By promoting formation of membrane microdomains essential for clustering of receptors and signalling platforms in the plasma membrane, ceramides accumulating as a result of membrane sphingomyelin breakdown are not only essential for assembly of signalling complexes and pathogen entry, but also act as signalling modulators, e. g. by regulating relay of phosphatidyl-inositol-3-kinase (PI3K) signalling. Their role in T lymphocyte functions has not been addressed as yet. We now show that measles virus (MV), which interacts with the surface of T cells and thereby efficiently interferes with stimulated dynamic reorganisation of their actin cytoskeleton, causes ceramide accumulation in human T cells in a neutral (NSM) and acid (ASM) sphingomyelinase-dependent manner. Ceramides induced by MV, but also bacterial sphingomyelinase, efficiently interfered with formation of membrane protrusions and T cell spreading and front/rear polarisation in response to beta1 integrin ligation or alphaCD3/CD28 activation, and this was rescued upon pharmacological or genetic ablation of ASM/NSM activity. Moreover, membrane ceramide accumulation downmodulated chemokine-induced T cell motility on fibronectin. Altogether, these findings highlight an as yet unrecognised concept of pathogens able to cause membrane ceramide accumulation to target essential processes in T cell activation and function by preventing stimulated actin cytoskeletal dynamics.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




Similar articles
-
Neutral sphingomyelinase in physiological and measles virus induced T cell suppression.PLoS Pathog. 2014 Dec 18;10(12):e1004574. doi: 10.1371/journal.ppat.1004574. eCollection 2014 Dec. PLoS Pathog. 2014. PMID: 25521388 Free PMC article.
-
Cytoskeletal dynamics: concepts in measles virus replication and immunomodulation.Viruses. 2011 Feb;3(2):102-117. doi: 10.3390/v3020102. Epub 2011 Jan 26. Viruses. 2011. PMID: 22049305 Free PMC article. Review.
-
DC-SIGN mediated sphingomyelinase-activation and ceramide generation is essential for enhancement of viral uptake in dendritic cells.PLoS Pathog. 2011 Feb;7(2):e1001290. doi: 10.1371/journal.ppat.1001290. Epub 2011 Feb 17. PLoS Pathog. 2011. PMID: 21379338 Free PMC article.
-
The role of sphingomyelin breakdown in measles virus immunmodulation.Cell Physiol Biochem. 2014;34(1):20-6. doi: 10.1159/000362981. Epub 2014 Jun 16. Cell Physiol Biochem. 2014. PMID: 24977478 Review.
-
The Neutral Sphingomyelinase 2 Is Required to Polarize and Sustain T Cell Receptor Signaling.Front Immunol. 2018 Apr 18;9:815. doi: 10.3389/fimmu.2018.00815. eCollection 2018. Front Immunol. 2018. PMID: 29720981 Free PMC article.
Cited by
-
Measles virus transmission from dendritic cells to T cells: formation of synapse-like interfaces concentrating viral and cellular components.J Virol. 2012 Sep;86(18):9773-81. doi: 10.1128/JVI.00458-12. Epub 2012 Jul 3. J Virol. 2012. PMID: 22761368 Free PMC article.
-
A Sphingosine 1-Phosphate Gradient Is Linked to the Cerebral Recruitment of T Helper and Regulatory T Helper Cells during Acute Ischemic Stroke.Int J Mol Sci. 2020 Aug 28;21(17):6242. doi: 10.3390/ijms21176242. Int J Mol Sci. 2020. PMID: 32872326 Free PMC article.
-
Neutral Sphingomyelinase-2 (NSM 2) Controls T Cell Metabolic Homeostasis and Reprogramming During Activation.Front Mol Biosci. 2020 Sep 4;7:217. doi: 10.3389/fmolb.2020.00217. eCollection 2020. Front Mol Biosci. 2020. PMID: 33088808 Free PMC article.
-
Neutral sphingomyelinase in physiological and measles virus induced T cell suppression.PLoS Pathog. 2014 Dec 18;10(12):e1004574. doi: 10.1371/journal.ppat.1004574. eCollection 2014 Dec. PLoS Pathog. 2014. PMID: 25521388 Free PMC article.
-
The Activity of the Neutral Sphingomyelinase Is Important in T Cell Recruitment and Directional Migration.Front Immunol. 2017 Aug 21;8:1007. doi: 10.3389/fimmu.2017.01007. eCollection 2017. Front Immunol. 2017. PMID: 28871263 Free PMC article.
References
-
- Bollinger CR, Teichgraber V, Gulbins E. Ceramide-enriched membrane domains. Biochim Biophys Acta. 2005;1746:284–294. - PubMed
-
- Grassme H, Jekle A, Riehle A, Schwarz H, Berger J, et al. CD95 signalling via ceramide-rich membrane rafts. J Biol Chem. 2001;276:20589–20596. - PubMed
-
- Church LD, Hessler G, Goodall JE, Rider DA, Workman CJ, et al. TNFR1-induced sphingomyelinase activation modulates TCR signalling by impairing store-operated Ca2+ influx. J Leukoc Biol. 2005;78:266–278. - PubMed
-
- Gulbins E, Grassme H. Ceramide and cell death receptor clustering. Biochim Biophys Acta. 2002;1585:139–145. - PubMed
-
- Abdel Shakor AB, Kwiatkowska K, Sobota A. Cell surface ceramide generation precedes and controls FcgammaRII clustering and phosphorylation in rafts. J Biol Chem. 2004;279:36778–36787. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
