

Chemokines and chemokine receptors: standing at the crossroads of immunobiology and neurobiology
- PMID: 19836265
- PMCID: PMC2787682
- DOI: 10.1016/j.immuni.2009.09.010
Chemokines and chemokine receptors: standing at the crossroads of immunobiology and neurobiology
Abstract
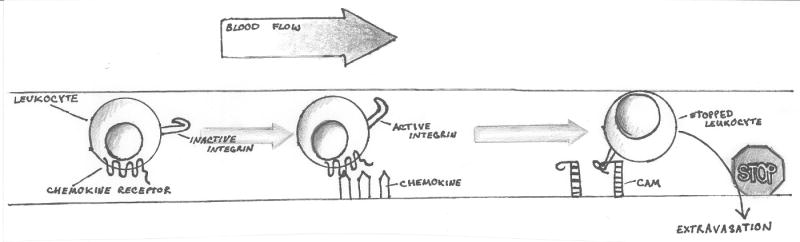
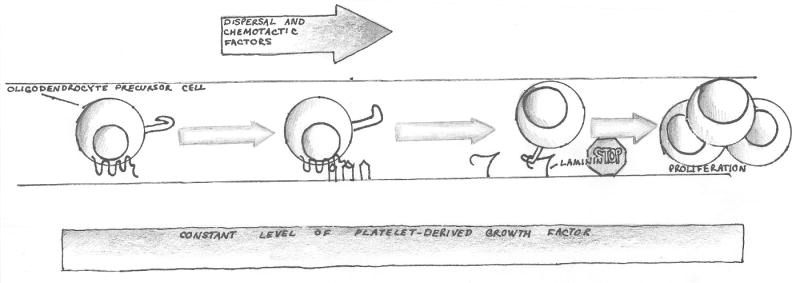
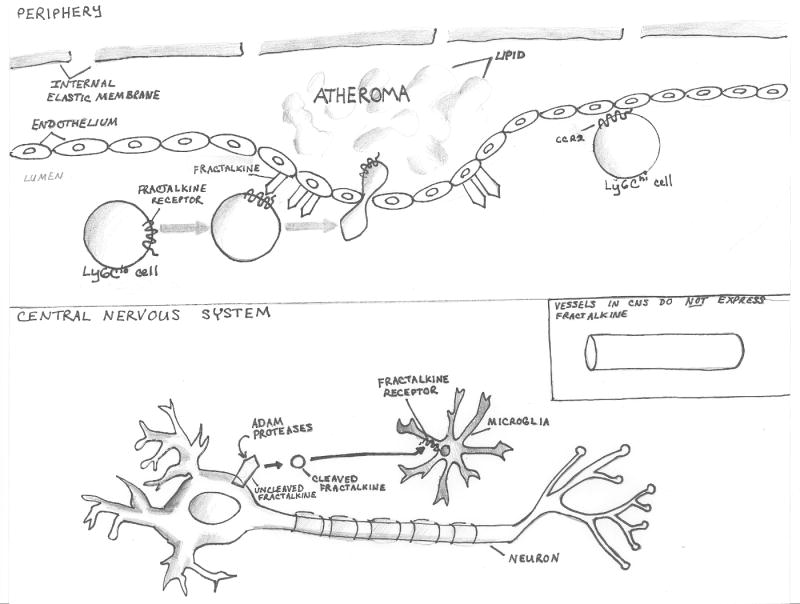
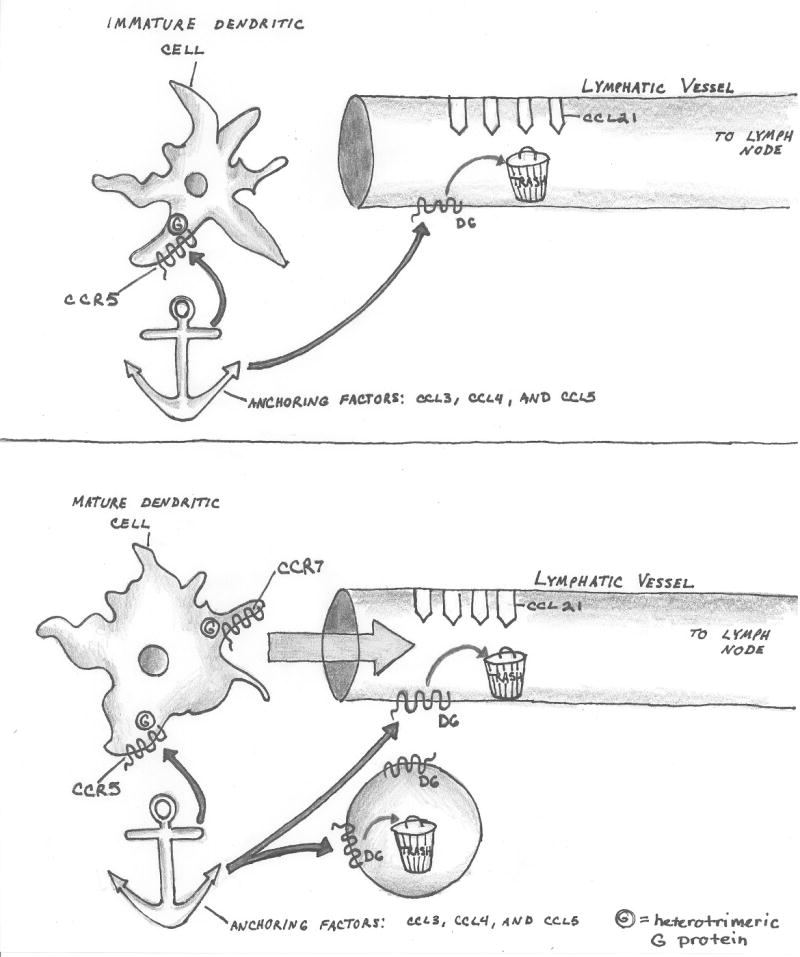
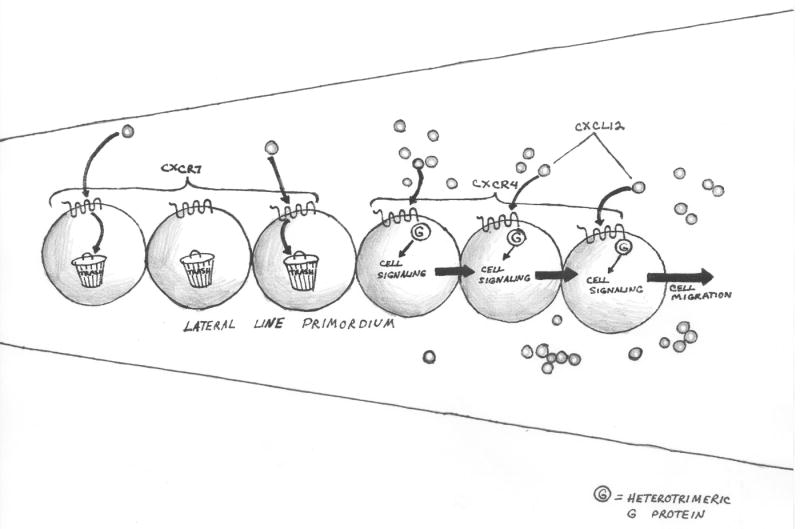
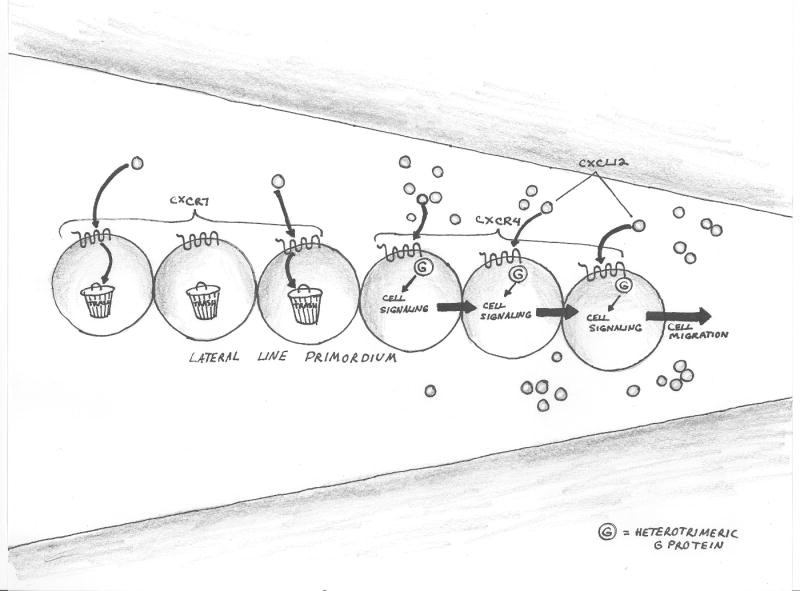
There are several molecular entities common to the immune and nervous systems. Salient among them are the chemokines and their receptors, which play remarkably varied and potent roles in immunobiology and neurobiology. This review limns several illustrative examples and presents general principles of chemokine action that are manifest in both systems. These include the following: (1) chemokines tend equally to arrest cells and to make them move, in the process of positioning target cells with spatiotemporal precision; (2) signaling and nonsignaling receptors collaborate to adjust the chemokine environment for maximal efficacy; and (3) expression of a single chemokine receptor on circulating blood cells and parenchymal cells is often used to coordinate complex tissue responses. The challenge is to integrate knowledge of the roles of key receptors (as well as their ligands) into a coherent account of events during pathologic processes, in order to guide therapeutic development.
Figures
References
-
- Abbadie C. Chemokines, chemokine receptors and pain. Trends Immunol. 2005;26:529–534. - PubMed
-
- Acosta JC, O'Loghlen A, Banito A, Guijarro MV, Augert A, Raguz S, Fumagalli M, Da CM, Brown C, Popov N, Takatsu Y, Melamed J, dda di FF, Bernard D, Hernando E, Gil J. Chemokine signaling via the CXCR2 receptor reinforces senescence. Cell. 2008;133:1006–1018. - PubMed
-
- Allen NJ, Attwell D. A chemokine-glutamate connection. Nat Neurosci. 2001;4:676–678. - PubMed
-
- Alon R, Feigelson S. From rolling to arrest on blood vessels: leukocyte tap dancing on endothelial integrin ligands and chemokines at sub-second contacts. Semin Immunol. 2002;14:93–104. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
