

Leptin and the systems neuroscience of meal size control
- PMID: 19836413
- PMCID: PMC2813996
- DOI: 10.1016/j.yfrne.2009.10.005
Leptin and the systems neuroscience of meal size control
Abstract
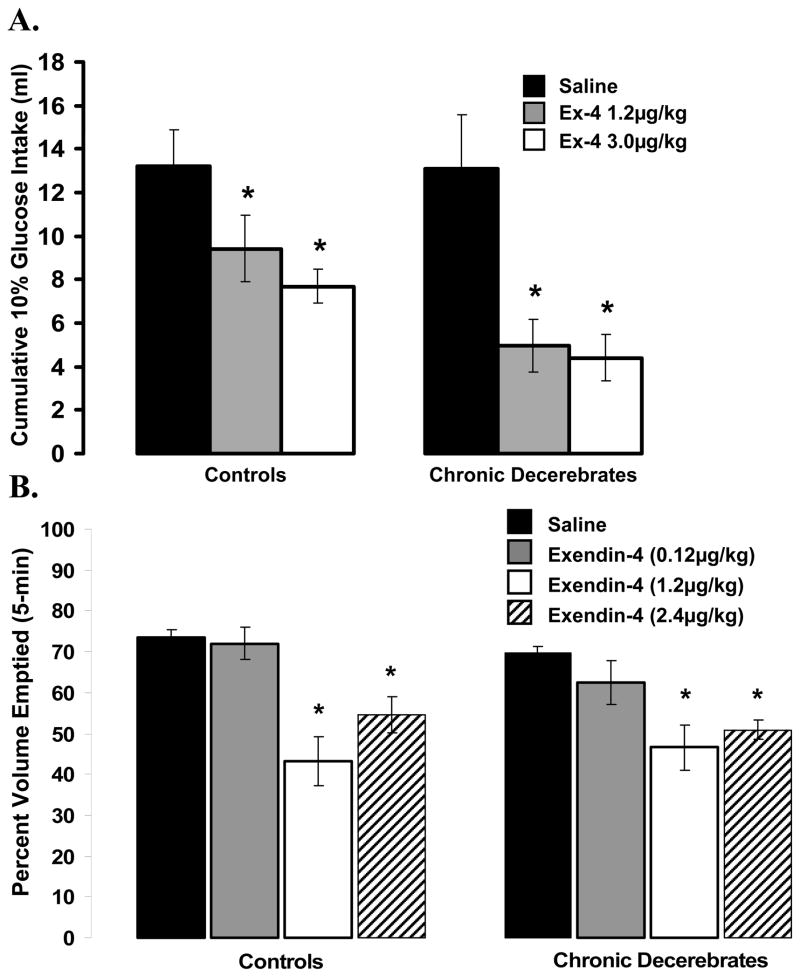
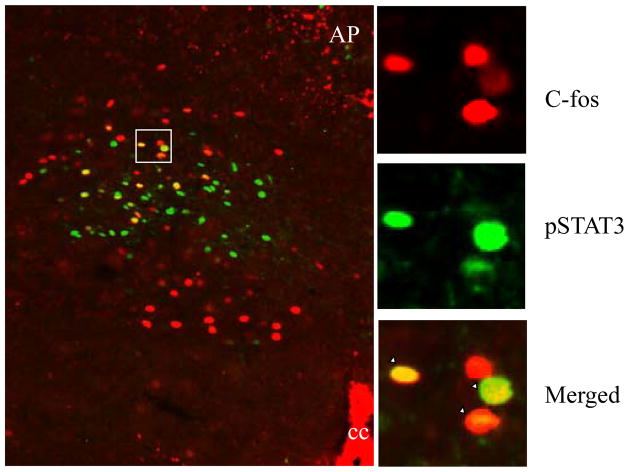
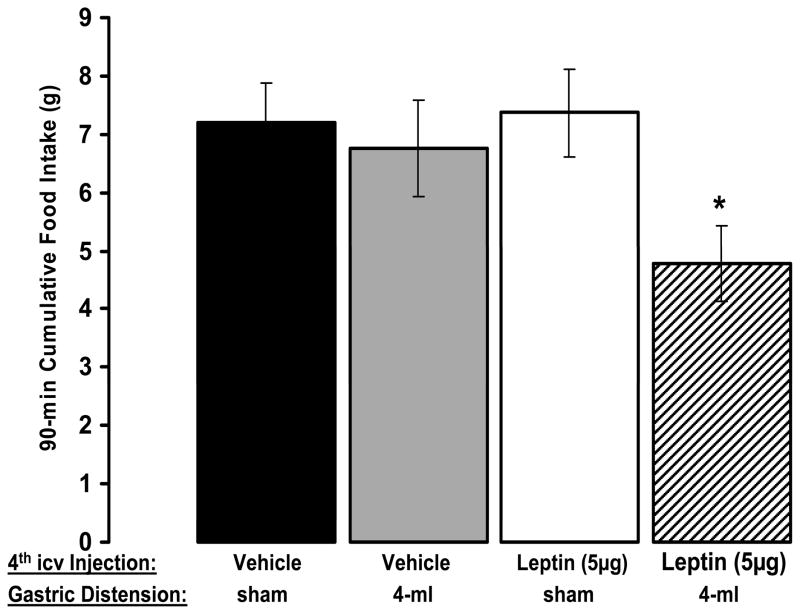
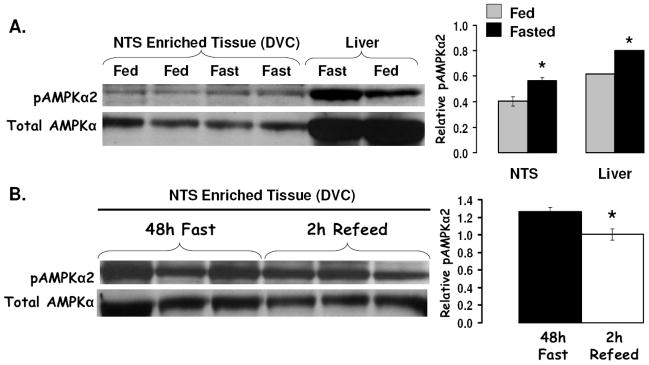
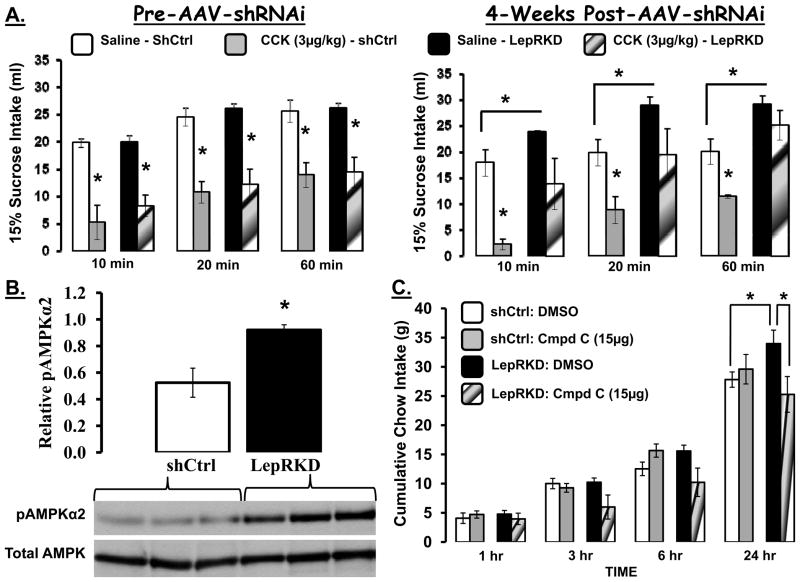
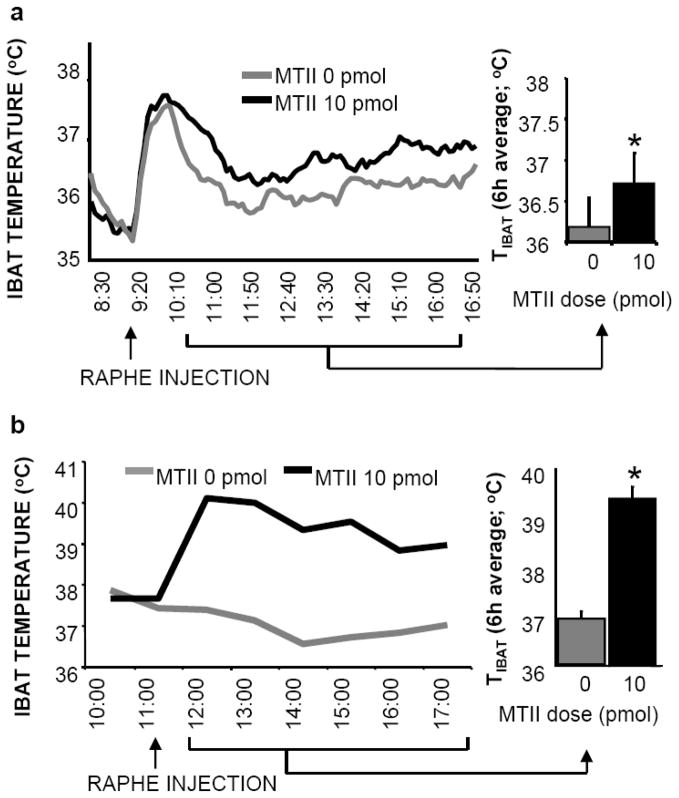
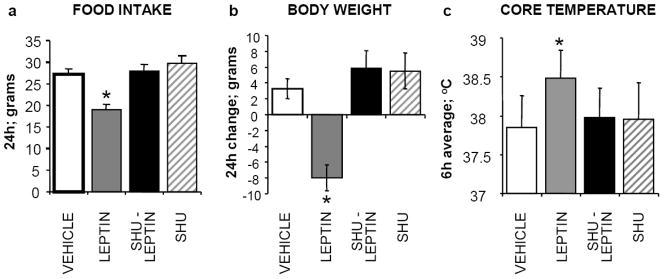
The development of effective pharmacotherapy for obesity will benefit from a more complete understanding of the neural pathways and the neurochemical signals whose actions result in the reduction of the size of meals. This review examines the neural control of meal size and the integration of two principal sources of that control--satiation signals arising from the gastrointestinal tract and CNS leptin signaling. Four types of integrations that are central to the control of meal size are described and each involves the neurons of the nucleus tractus solitarius (NTS) in the dorsal hindbrain. Data discussed show that NTS neurons integrate information arising from: (1) ascending GI-derived vagal afferent projections, (2) descending neuropeptidergic projections from leptin-activated arcuate and paraventricular nucleus neurons, (3) leptin signaling in NTS neurons themselves and (4) melanocortinergic projections from NTS and hypothalamic POMC neurons to NTS neurons and melanocortinergic modulation of vagal afferent nerve terminals that are presynaptic to NTS neurons.
2009 Elsevier Inc. All rights reserved.
Figures
References
-
- Rolls BJ, Drewnowski A, Ledikwe JH. Changing the energy density of the diet as a strategy for weight management. J Am Diet Assoc. 2005;105:S98–103. - PubMed
-
- Bjorntorp P. Thrifty genes and human obesity. Are we chasing ghosts? Lancet. 2001;358:1006–1008. - PubMed
-
- Hayes M, Chustek M, Heshka S, Wang Z, Pietrobelli A, Heymsfield SB. Low physical activity levels of modern Homo sapiens among free-ranging mammals. Int J Obes (Lond) 2005;29:151–156. - PubMed
-
- Leonard WR. Food for thought. Dietary change was a driving force in human evolution. Sci Am. 2002;287:106–115. - PubMed
-
- Leonard WR, Robertson ML. Nutritional requirements and human evolution: a bioenergetics model. Am J Hum Biol. 1992;4:179–195. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
