

Review
doi: 10.1016/j.cell.2009.09.029.
Phototransduction motifs and variations
Affiliations
- PMID: 19837030
- PMCID: PMC2885920
- DOI: 10.1016/j.cell.2009.09.029
Item in Clipboard
Review
Phototransduction motifs and variations
Cell.
.
Abstract
Seeing begins in the photoreceptors, where light is absorbed and signaled to the nervous system. Throughout the animal kingdom, photoreceptors are diverse in design and purpose. Nonetheless, phototransduction-the mechanism by which absorbed photons are converted into an electrical response-is highly conserved and based almost exclusively on a single class of photoproteins, the opsins. In this Review, we survey the G protein-coupled signaling cascades downstream from opsins in photoreceptors across vertebrate and invertebrate species, noting their similarities as well as differences.
Figures
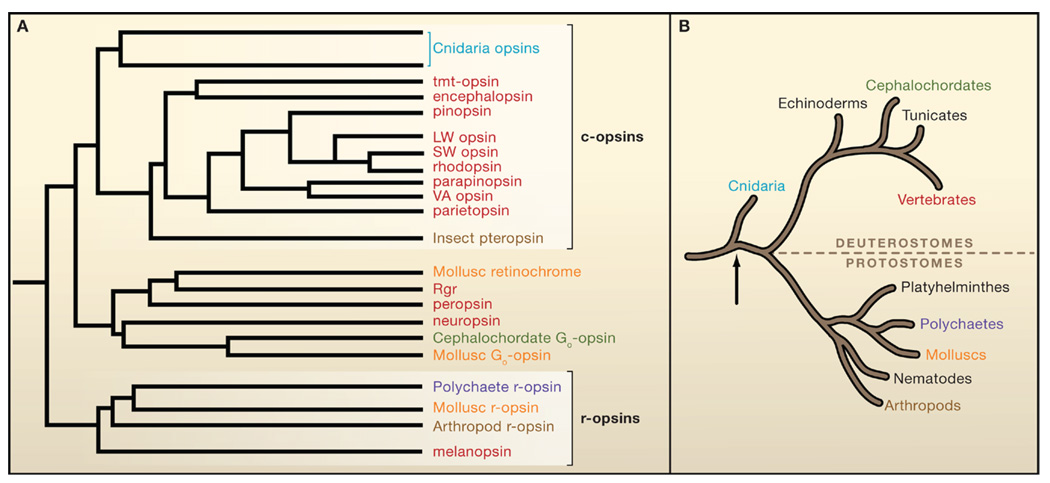
(A) Schematic phylogenetic tree of the opsin family. Depicted opsins (vertebrate unless otherwise stated) are color-coded with respect to the phyla of origin shown in (B). There are two main groups: c-opsins and r-opsins, together with a miscellaneous group (Go-opsin, etc.) more closely related to c-opsins than r-opsins. Branch lengths are arbitrary. Simplified from that originally derived with a maximum-likelihood algorithm by Suga et al. (2008). (B) Simplified evolutionary tree of present-day animal phyla. Modern-day bilaterians comprise the protostome invertebrates and the deuterostomes, which include the vertebrate lineage. These two lines diverged ~550 million years ago (Mya), by which time an “Urbilaterian,” a common ancestor, had already evolved with both rhabdomeric- and ciliary-type photoreceptors (Arendt, 2003). Arrow indicates the Cnidarian (prebilaterians)/bilaterian split.
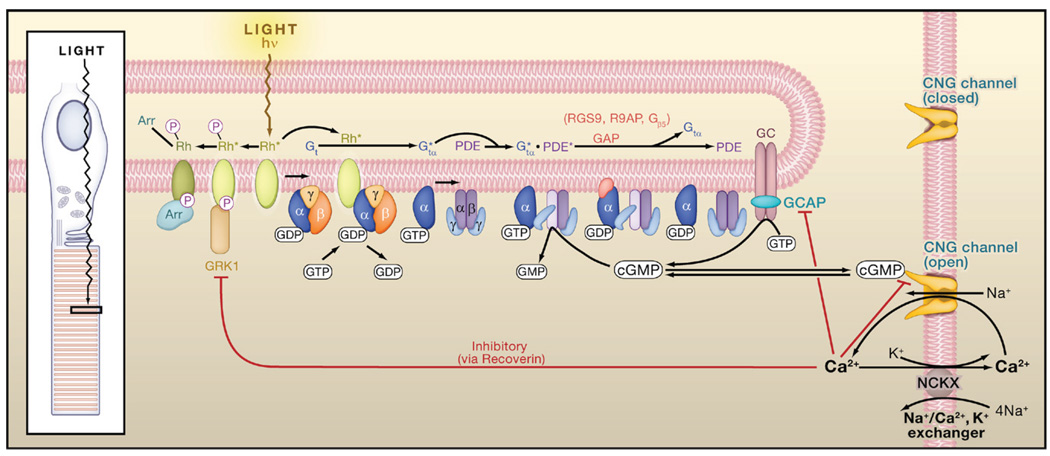
Light converts rhodopsin into an active form, Rh*, which activates heterotrimeric Gt by GTP-GDP exchange. Active Gtα (Gtα*) binds to and activates phosphodiesterase (PDE), which hydrolyzes cyclic GMP (cGMP) to GMP, thereby closing the cyclic-nucleotide-gated (CNG) channels that are open in darkness. hν, photon; Rh*-P, phosphorylated Rh*, which retains partial Rh* activity; Rh*-P-Arr, phosphorylated Rh* with arrestin bound, rendered fully inactive. Red lines ending in a small bar indicate negative-feedback (inhibitory) pathways via Ca2+. The inhibition by Ca2+ on the cGMP gating of the CNG channel is via a Ca2+-binding protein that may be calmodulin. Based on Yau (1994), Luo et al. (2008b), and Pugh et al. (1999). Inset: schematic diagram of the ciliary rod photoreceptor, with a light-sensitive outer segment formed from a highly expanded cilium.
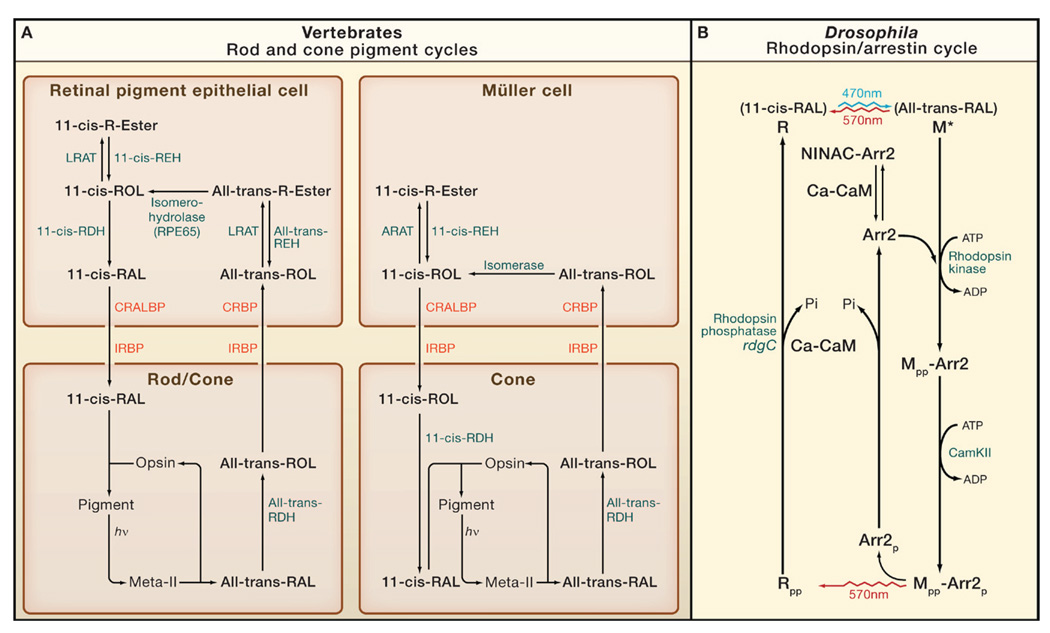
(A) Rods and cones. There are two cycles: one involving the retinal pigment epithelial cell and used by both rods and cones, and the other involving the Müller cell and used exclusively by cones (Travis et al., 2007 and Muniz et al., 2007). hν, photon; RAL, retinal; ROL, retinol; R-ester, retinyl ester; IRBP, interphotoreceptor retinoid-binding protein; CRBP, cellular retinol-binding protein; CRALBP, cellular retinaldehyde-binding protein; RDH, retinol dehydrogenase; REH, retinyl ester hydrolase; LRAT, lecithin:retinol acyl transferase; ARAT, acyl-CoA:retinol acyl transferase. LRAT and RPE65 are absent in Müller cells. The isomerase and ARAT in Müller cells are kinetically coupled and together named isomerosynthase. (B) Drosophila photoreceptor. Blue light (470 nm) photoisomerizes 11-cis-3-hydroxy-retinal in rhodopsin (R) to all-trans 3-hydroxy-retinal (top), generating active metarhodopsin (M*). Photoregeneration is achieved simply by long-wavelength light (570 nm), which reisomerizes all-trans to 11-cis, thereby reconverting M to R irrespective of whether it is phosphorylated (indicated by “-pp”) or bound to arrestin (Arr2). M* is thermostable and continues to activate Gq until it binds Arr2. M is also phosphorylated by rhodopsin kinase (RK) on C-terminal serines, but this is not required for Arr2 binding or response termination. The Mpp-Arr2 state is a target for clathrin-mediated endocytosis, but this endocytosis is inhibited by the CaMKII-dependent phosphorylation of Arr2. After Mpp is photoreconverted to Rpp, Arr2 is released, as long as it has been phosphorylated (Arr2p). Rpp is dephosphorylated by Ca-CaM-dependent rhodopsin phosphatase (coded by rdgC) to recreate the ground state, R. Prior to Ca2+ influx, Arr2 is prevented from binding to M*, by being bound to NINAC or a NINAC-regulated target. Once channels open to allow Ca2+ influx, Ca-CaM releases Arr2 to allow it to rapidly bind to M* (Liu et al., 2008).
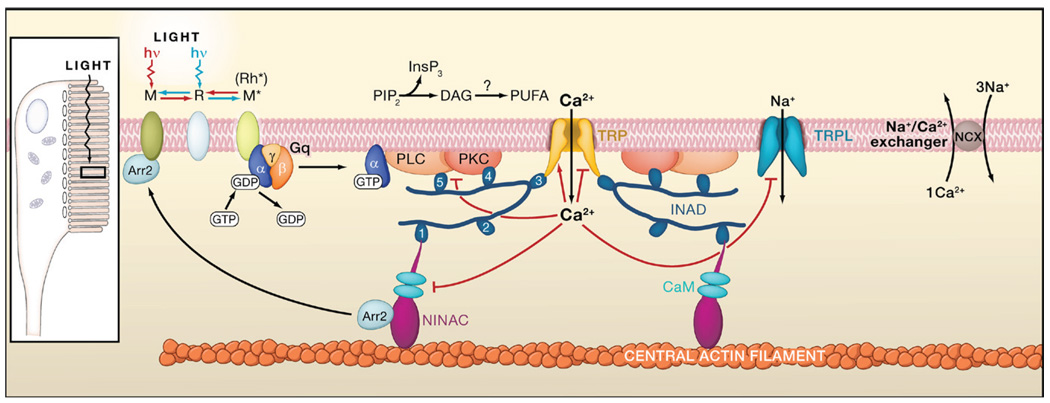
Absorption of a photon by rhodopsin (R) converts it to the thermostable, active metarhodopsin state (M* or Rh*), which activates heterotrimeric Gq by GTP-GDP exchange essentially the same as in vertebrate rods. Active Gαq binds to and activates phospholipase C (PLC), which hydrolyzes phosphatidylinositol 4,5-bisphosphate (PIP2) to inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG), with the latter potentially producing polyunsaturated fatty acids (PUFAs) via a DAG lipase. Two classes of light-sensitive channels (TRP and TRPL, with the first being primarily Ca2+ permeable) are activated by a still-unknown membrane-delimited effect of PLC activity. Ca2+ influx feeds back positively and negatively at multiple sites (indicated by red lines ending in arrowheads and small bars, respectively), including PKC (required for inactivation of PLC), NINAC/arrestin (Arr2), and the TRP/TRPL channels. Ca2+ is extruded by a Na/Ca exchanger. TRP, PKC, and PLC are assembled into a signaling complex by the scaffolding protein INAD, possibly linked to the F-actin core via NINAC, a CaM-binding class III myosin. INAD has 5 PDZ domains, associated preferentially with different targets. The precise composition of the native complex is uncertain. Inset: schematic diagram of the rhabdomeric Drosophila photoreceptor, with microvilli forming a light-guiding rhabdomere. Submicrovillar cisternae at 10–100 nm beneath the base of the microvilli may release Ca2+ via InsP3 receptors in many rhabdomeric photoreceptors. However, InsP3 appears to play no role in photoactivation in Drosophila.
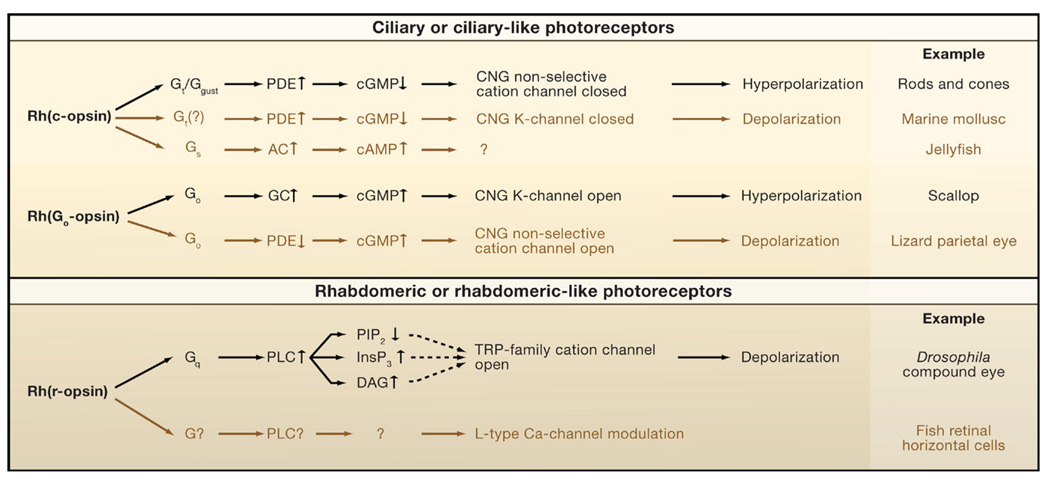
There are two primary motifs, one mediated by cyclic nucleotides and the other by phospholipase C (PLC), segregated in ciliary and rhabdomeric photoreceptors, respectively. Within each primary motif, the canonical pathways are shown in black, and the noncanonical ones in brown to indicate their rare occurrence as currently known. The cyclic-nucleotide motif has two submotifs, mediated by Gt (or its close homolog, Ggust) and Go. In the Gt (?) pathway, found in the abdominal ganglion of the marine slug, Onchidium, the involvement of Gt is by inference only, without molecular identity, hence the question mark. The Gs pathway, reported in a box jellyfish, is tentative at present. The second, noncanonical Go pathway, found in the vertebrate parietal-eye photoreceptor, differs from the canonical Go pathway by involving a decrease in phosphodiesterase (PDE) activity rather than an increase in guanylate cyclase (GC) activity, with the same end result, namely, an increase in cyclic GMP (cGMP). The Go-PDE pathway may be unique to the parietal-eye photoreceptor, called for by the chromatic antagonism in this cell. In the PLC canonical pathway, the dashed lines indicate that the channel-activating messenger is still unclear. PLC depletes phosphatidylinositol 4,5-bisphosphate (PIP2) by hydrolyzing it into diacylglycerol (DAG) and inositol 1,4,5-trisphosphate (InsP3) with the latter releasing Ca2+ from intracellular stores; there is evidence implicating PIP2, DAG, InsP3 (via intracellular Ca2+ release), and a metabolite of DAG in the gating of the TRP-family nonselective cation channels. Ca2+ may also act synergistically with lipid messengers rather than as an activator in its own right. The canonical Gq pathway is typified by Drosophila and Limulus photoreceptors. The noncanonical Gq pathway is tentative, found in vertebrate intrinsically photosensitive horizontal cells (ipHCs), and suggested by current evidence to use melanopsin (an r-opsin) as pigment; however, there is no information on the intermediate steps or their molecular components. There are other differences between motifs or variations within a motif. For example, prominent negative feedbacks mediated by Ca2+ exist in the canonical Gt-mediated pathway but are apparently absent in the canonical Go-mediated pathway.
References
-
- Arendt D. Evolution of eyes and photoreceptor cell types. Int. J. Dev. Biol. 2003;47:563–571. - PubMed
-
- Arshavsky VY, Lamb TD, Pugh EN., Jr G Proteins and phototransduction. Annu. Rev. Physiol. 2002;64:153–187. - PubMed
-
- Ashmore LJ, Sehgal A. A fly’s eye view of circadian entrainment. J. Biol. Rhythms. 2003;18:206–216. - PubMed
-
- Bandyopadhyay BC, Payne R. Variants of TRP ion channel mRNA present in horseshoe crab ventral eye and brain. J. Neurochem. 2004;91:825–835. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 EY006837/EY/NEI NIH HHS/United States
- R01 EY014596/EY/NEI NIH HHS/United States
- BB/G006865/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/D007585/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- R01 DC006904/DC/NIDCD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
