

Norrin, frizzled-4, and Lrp5 signaling in endothelial cells controls a genetic program for retinal vascularization
- PMID: 19837032
- PMCID: PMC2779707
- DOI: 10.1016/j.cell.2009.07.047
Norrin, frizzled-4, and Lrp5 signaling in endothelial cells controls a genetic program for retinal vascularization
Erratum in
- Cell. 2010 Apr 2;141(1):191
Abstract
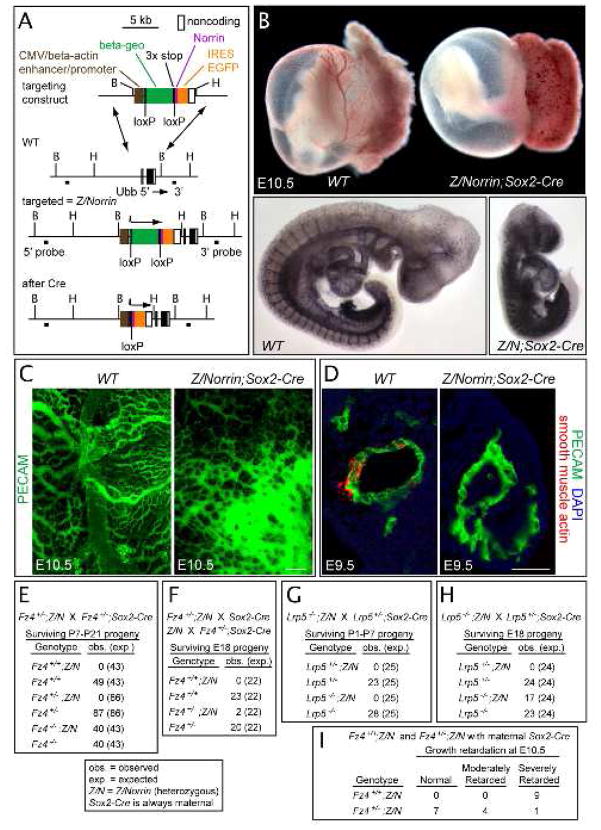
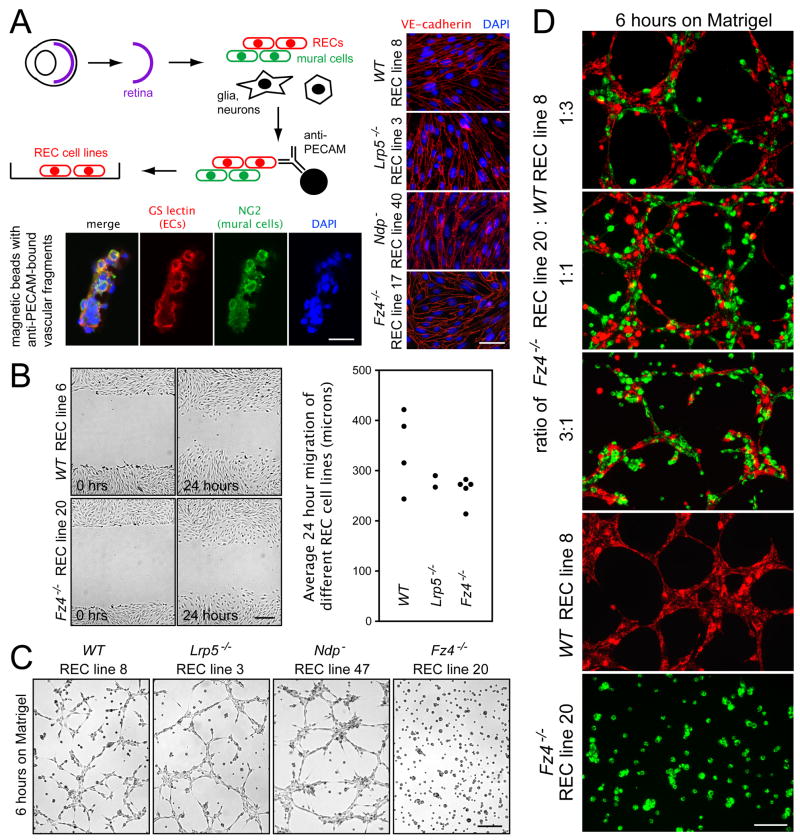
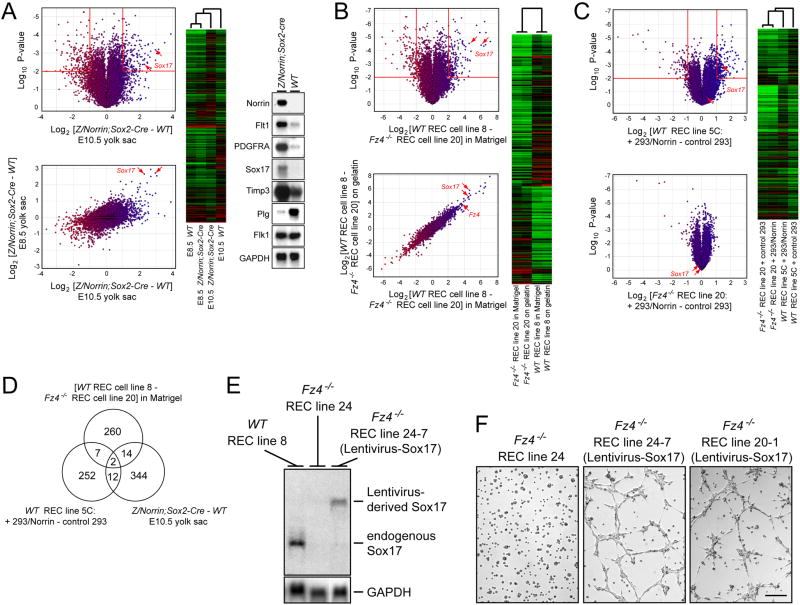
Disorders of vascular structure and function play a central role in a wide variety of CNS diseases. Mutations in the Frizzled-4 (Fz4) receptor, Lrp5 coreceptor, or Norrin ligand cause retinal hypovascularization, but the mechanisms by which Norrin/Fz4/Lrp signaling controls vascular development have not been defined. Using mouse genetic and cell culture models, we show that loss of Fz4 signaling in endothelial cells causes defective vascular growth, which leads to chronic but reversible silencing of retinal neurons. Loss of Fz4 in all endothelial cells disrupts the blood brain barrier in the cerebellum, whereas excessive Fz4 signaling disrupts embryonic angiogenesis. Sox17, a transcription factor that is upregulated by Norrin/Fz4/Lrp signaling, plays a central role in inducing the angiogenic program controlled by Norrin/Fz4/Lrp. These experiments establish a cellular basis for retinal hypovascularization diseases due to insufficient Frizzled signaling, and they suggest a broader role for Frizzled signaling in vascular growth, remodeling, maintenance, and disease.
Figures
Comment in
-
Eyeing up new Wnt pathway players.Cell. 2009 Oct 16;139(2):227-9. doi: 10.1016/j.cell.2009.09.027. Cell. 2009. PMID: 19837026
References
-
- Adams RH, Alitalo K. Molecular regulation of angiogenesis and lymphangiogenesis. Nat Rev Mol Cell Biol. 2007;8:464–478. - PubMed
-
- Alm A. Ocular circulation. In: Hart WM, editor. Adler’s Physiology of the Eye. 9. Saint Louis: Mosby; 1992. pp. 198–227.
-
- Armulik A, Abramsson A, Betsholtz C. Endothelial/pericyte interactions. Circ Res. 2005;97:512–523. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
