

Complement protein C1q reduces the stoichiometric threshold for antibody-mediated neutralization of West Nile virus
- PMID: 19837377
- PMCID: PMC2782387
- DOI: 10.1016/j.chom.2009.09.003
Complement protein C1q reduces the stoichiometric threshold for antibody-mediated neutralization of West Nile virus
Abstract
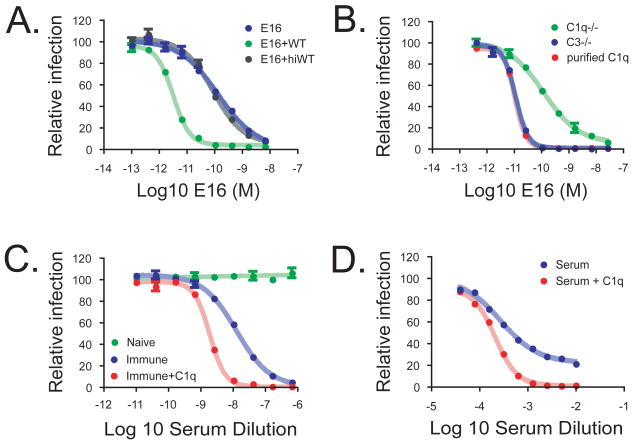
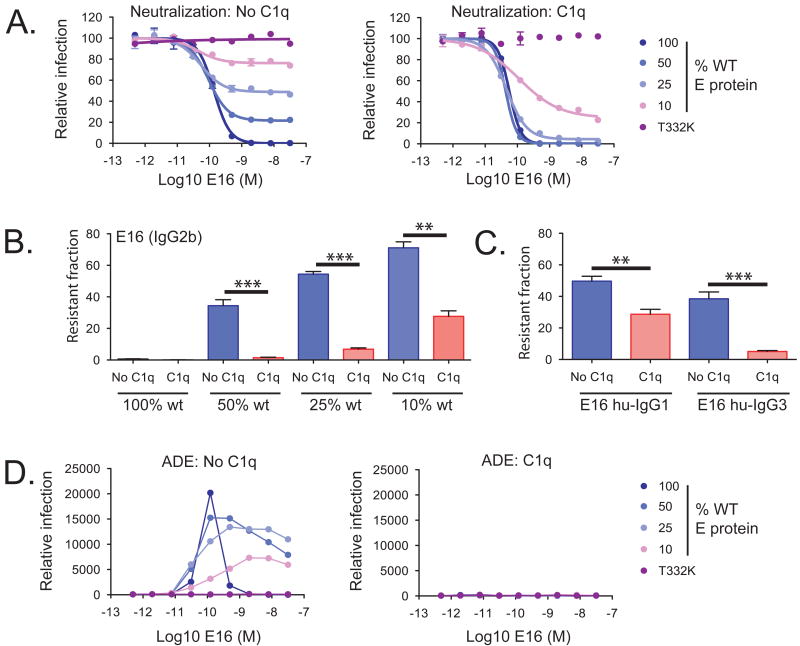
Virus neutralization is governed by the number of antibodies that bind a virion during the cellular entry process. Cellular and serum factors that interact with antibodies have the potential to modulate neutralization potency. Although the addition of serum complement can increase the neutralizing activity of antiviral antibodies in vitro, the mechanism and significance of this augmented potency in vivo remain uncertain. Herein, we show that the complement component C1q increases the potency of antibodies against West Nile virus by modulating the stoichiometric requirements for neutralization. The addition of C1q does not result in virolysis but instead reduces the number of antibodies that must bind the virion to neutralize infectivity. For IgG subclasses that bind C1q avidly, this reduced stoichiometric threshold falls below the minimal number of antibodies required for antibody-dependent enhancement (ADE) of infection of cells expressing Fc-gamma receptors (CD32) and explains how C1q restricts the ADE of flavivirus infection.
Conflict of interest statement
Figures



Similar articles
-
Complement protein C1q inhibits antibody-dependent enhancement of flavivirus infection in an IgG subclass-specific manner.Cell Host Microbe. 2007 Dec 13;2(6):417-26. doi: 10.1016/j.chom.2007.09.015. Cell Host Microbe. 2007. PMID: 18078693 Free PMC article.
-
A plant-produced vaccine protects mice against lethal West Nile virus infection without enhancing Zika or dengue virus infectivity.Vaccine. 2018 Mar 27;36(14):1846-1852. doi: 10.1016/j.vaccine.2018.02.073. Epub 2018 Feb 26. Vaccine. 2018. PMID: 29490880 Free PMC article.
-
A dynamic landscape for antibody binding modulates antibody-mediated neutralization of West Nile virus.PLoS Pathog. 2011 Jun;7(6):e1002111. doi: 10.1371/journal.ppat.1002111. Epub 2011 Jun 30. PLoS Pathog. 2011. PMID: 21738473 Free PMC article.
-
The molecular basis of antibody-mediated neutralization of West Nile virus.Expert Opin Biol Ther. 2007 Jun;7(6):885-92. doi: 10.1517/14712598.7.6.885. Expert Opin Biol Ther. 2007. PMID: 17555373 Review.
-
Antibody-Dependent Enhancement (ADE) and the role of complement system in disease pathogenesis.Mol Immunol. 2022 Dec;152:172-182. doi: 10.1016/j.molimm.2022.11.010. Epub 2022 Nov 10. Mol Immunol. 2022. PMID: 36371813 Free PMC article. Review.
Cited by
-
Serum peptidome profiles immune response of COVID-19 Vaccine administration.Front Immunol. 2022 Aug 24;13:956369. doi: 10.3389/fimmu.2022.956369. eCollection 2022. Front Immunol. 2022. PMID: 36091008 Free PMC article.
-
Complement in malaria immunity and vaccines.Immunol Rev. 2020 Jan;293(1):38-56. doi: 10.1111/imr.12802. Epub 2019 Sep 26. Immunol Rev. 2020. PMID: 31556468 Free PMC article. Review.
-
Analysis of Complement-Mediated Lysis of Simian Immunodeficiency Virus (SIV) and SIV-Infected Cells Reveals Sex Differences in Vaccine-Induced Immune Responses in Rhesus Macaques.J Virol. 2018 Sep 12;92(19):e00721-18. doi: 10.1128/JVI.00721-18. Print 2018 Oct 1. J Virol. 2018. PMID: 30021899 Free PMC article.
-
The neutralizing capacity of antibodies elicited by parainfluenza virus infection of African Green Monkeys is dependent on complement.Virology. 2014 Jul;460-461:23-33. doi: 10.1016/j.virol.2014.05.004. Epub 2014 May 29. Virology. 2014. PMID: 25010267 Free PMC article.
-
Complement-mediated enhancement of SARS-CoV-2 antibody neutralisation potency in vaccinated individuals.Nat Commun. 2025 Mar 18;16(1):2666. doi: 10.1038/s41467-025-57947-8. Nat Commun. 2025. PMID: 40102474 Free PMC article.
References
-
- Andrewes CH, Elford WJ. Observations on anti-phage sera. I. ‘The percentage law’. British Journal of Experimental Pathology. 1933;14:367–376.
-
- Ben-Nathan D, Lustig S, Tam G, Robinzon S, Segal S, Rager-Zisman B. Prophylactic and therapeutic efficacy of human intravenous immunoglobulin in treating west nile virus infection in mice. The Journal of infectious diseases. 2003;188:5–12. - PubMed
-
- Burnet FM, Keogh EV, Lush D. The immunological reactions of the filterable viruses. The Australian Journal of Experimental Biology and Medical Science. 1937;15:227–368.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
