

PPAR-delta senses and orchestrates clearance of apoptotic cells to promote tolerance
- PMID: 19838202
- PMCID: PMC2783696
- DOI: 10.1038/nm.2048
PPAR-delta senses and orchestrates clearance of apoptotic cells to promote tolerance
Abstract
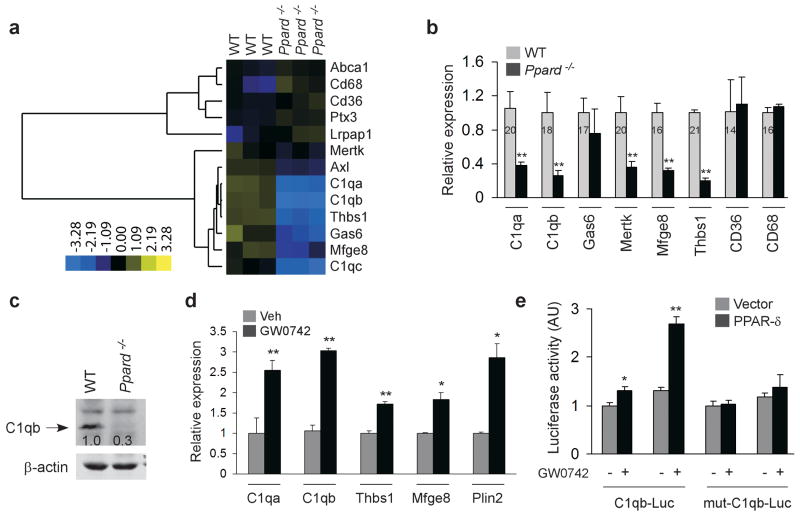
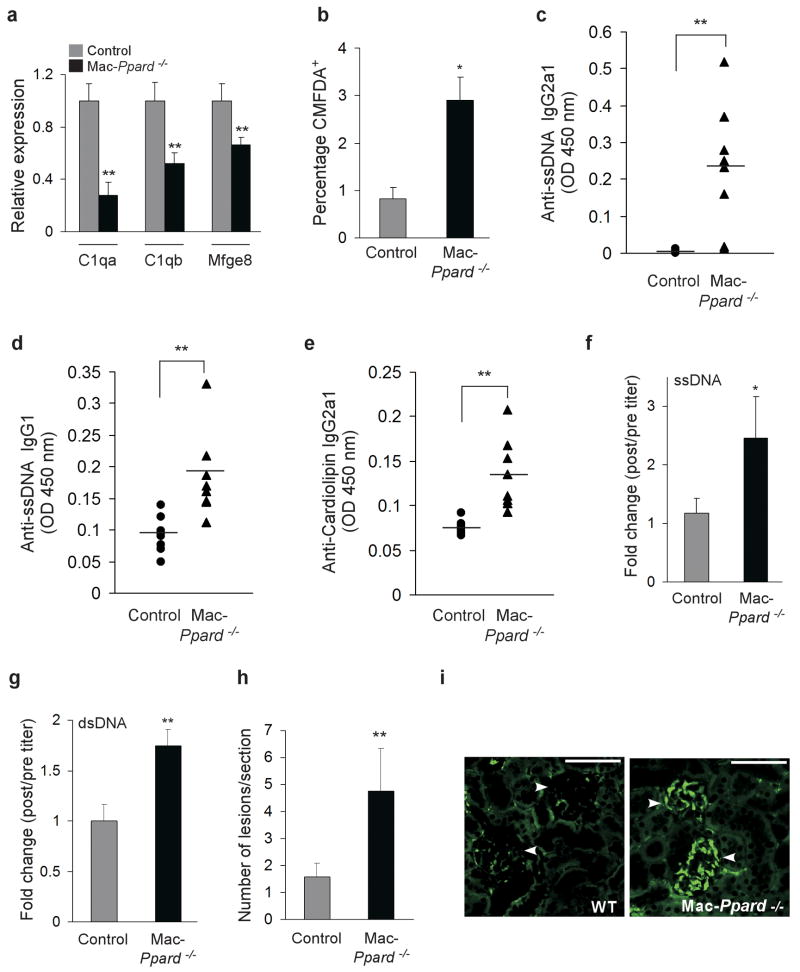
Macrophages rapidly engulf apoptotic cells to limit the release of noxious cellular contents and to restrict autoimmune responses against self antigens. Although factors participating in recognition and engulfment of apoptotic cells have been identified, the transcriptional basis for the sensing and the silent disposal of apoptotic cells is unknown. Here we show that peroxisome proliferator-activated receptor-delta (PPAR-delta) is induced when macrophages engulf apoptotic cells and functions as a transcriptional sensor of dying cells. Genetic deletion of PPAR-delta decreases expression of opsonins such as complement component-1qb (C1qb), resulting in impairment of apoptotic cell clearance and reduction in anti-inflammatory cytokine production. This increases autoantibody production and predisposes global and macrophage-specific Ppard(-/-) mice to autoimmune kidney disease, a phenotype resembling the human disease systemic lupus erythematosus. Thus, PPAR-delta has a pivotal role in orchestrating the timely disposal of apoptotic cells by macrophages, ensuring that tolerance to self is maintained.
Figures




Comment in
-
Autoimmunity: apoptotic fats grease transcription.Nat Med. 2009 Nov;15(11):1246-8. doi: 10.1038/nm1109-1246. Nat Med. 2009. PMID: 19893553 No abstract available.
References
-
- Savill J, Fadok V. Corpse clearance defines the meaning of cell death. Nature. 2000;407:784–8. - PubMed
-
- Henson PM, Bratton DL, Fadok VA. The phosphatidylserine receptor: a crucial molecular switch? Nat Rev Mol Cell Biol. 2001;2:627–33. - PubMed
-
- Savill J, Dransfield I, Gregory C, Haslett C. A blast from the past: clearance of apoptotic cells regulates immune responses. Nat Rev Immunol. 2002;2:965–75. - PubMed
-
- Lauber K, Blumenthal SG, Waibel M, Wesselborg S. Clearance of apoptotic cells: getting rid of the corpses. Mol Cell. 2004;14:277–87. - PubMed
-
- Erwig LP, Henson PM. Clearance of apoptotic cells by phagocytes. Cell Death Differ. 2008;15:243–50. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- DK062386/DK/NIDDK NIH HHS/United States
- HL076746/HL/NHLBI NIH HHS/United States
- R01 DK081405/DK/NIDDK NIH HHS/United States
- K08 DK062386/DK/NIDDK NIH HHS/United States
- AI066402/AI/NIAID NIH HHS/United States
- DK67592/DK/NIDDK NIH HHS/United States
- F31 AI066402/AI/NIAID NIH HHS/United States
- DK081405/DK/NIDDK NIH HHS/United States
- R01 DE014385/DE/NIDCR NIH HHS/United States
- DE14385/DE/NIDCR NIH HHS/United States
- R01 DK067592/DK/NIDDK NIH HHS/United States
- R01 DK076760/DK/NIDDK NIH HHS/United States
- T32 AI007290/AI/NIAID NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- R01 HL076746/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
