

Animal models for Alzheimer's disease and frontotemporal dementia: a perspective
- PMID: 19839939
- PMCID: PMC2785514
- DOI: 10.1042/AN20090042
Animal models for Alzheimer's disease and frontotemporal dementia: a perspective
Abstract
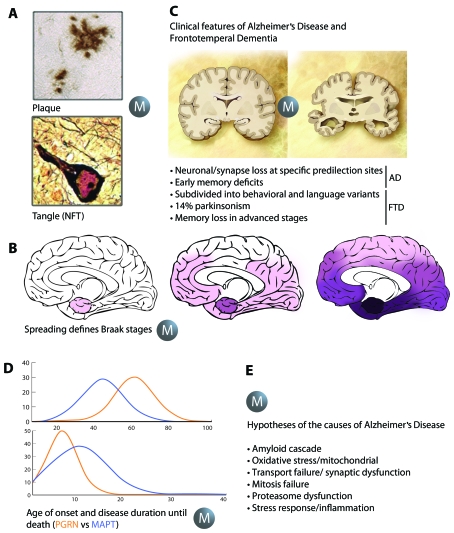
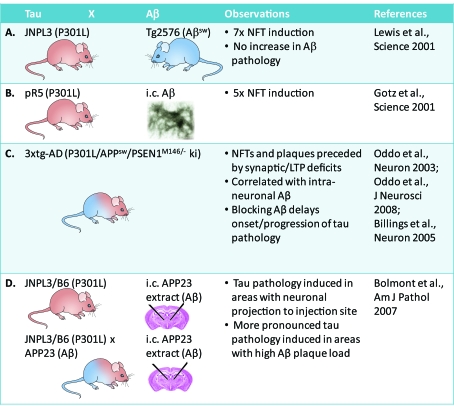
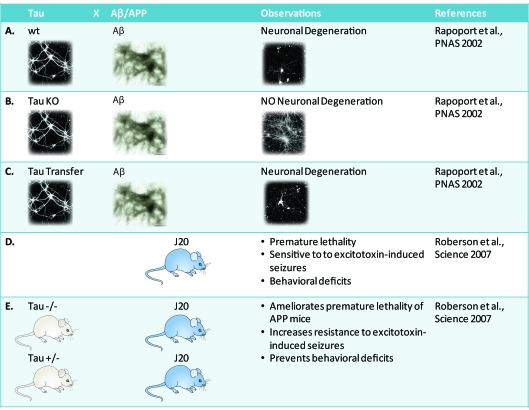
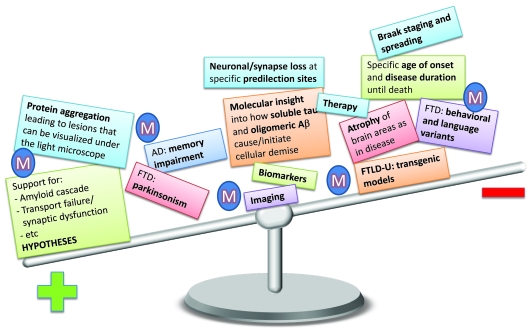
In dementia research, animal models have become indispensable tools. They not only model aspects of the human condition, but also simulate processes that occur in humans and hence provide insight into how disease is initiated and propagated. The present review discusses two prominent human neurodegenerative disorders, Alzheimer's disease and frontotemporal dementia. It discusses what we would like to model in animals and highlights some of the more recent achievements using species as diverse as mice, fish, flies and worms. Advances in imaging and therapy are explored. We also discuss some anticipated new models and developments. These will reveal how key players in the pathogenesis of Alzheimer's disease and frontotemporal dementia, such as the peptide Aβ (amyloid β) and the protein tau, cause neuronal dysfunction and eventually, neuronal demise. Understanding these processes fully will lead to early diagnosis and therapy.
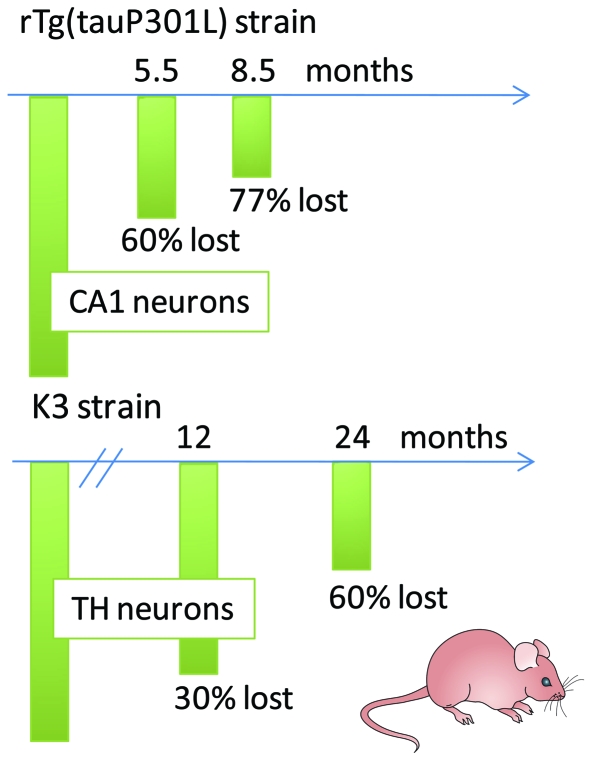
Figures
References
-
- Allen B, Ingram E, Takao M, Smith MJ, Jakes R, Virdee K, Yoshida H, Holzer M, Craxton M, Emson PC, Atzori C, Migheli A, Crowther RA, Ghetti B, Spillantini MG, Goedert M. Abundant tau filaments and nonapoptotic neurodegeneration in transgenic mice expressing human P301S tau protein. J Neurosci. 2002;22:9340–9351. - PMC - PubMed
-
- Andorfer C, Kress Y, Espinoza M, de Silva R, Tucker KL, Barde YA, Duff K, Davies P. Hyperphosphorylation and aggregation of tau in mice expressing normal human tau isoforms. J Neurochem. 2003;86:582–590. - PubMed
-
- Arnold SE, Hyman BT, Flory J, Damasio AR, Van Hoesen GW. The topographical and neuroanatomical distribution of neurofibrillary tangles and neuritic plaques in the cerebral cortex of patients with Alzheimer's disease. Cereb Cortex. 1991;1:103–116. - PubMed
-
- Bach P, Tschape JA, Kopietz F, Braun G, Baade JK, Wiederhold KH, Staufenbiel M, Prinz M, Deller T, Kalinke U, Buchholz CJ, Muller UC. Vaccination with Aβ-displaying virus-like particles reduces soluble and insoluble cerebral Aβ and lowers plaque burden in APP transgenic mice. J Immunol. 2009;182:7613–7624. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources