

Mechanical loading regulates NFATc1 and beta-catenin signaling through a GSK3beta control node
- PMID: 19840939
- PMCID: PMC2787323
- DOI: 10.1074/jbc.M109.039453
Mechanical loading regulates NFATc1 and beta-catenin signaling through a GSK3beta control node
Abstract
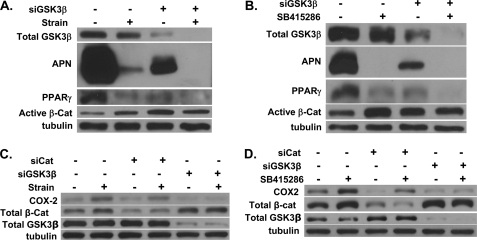
Mechanical stimulation can prevent adipogenic and improve osteogenic lineage allocation of mesenchymal stem cells (MSC), an effect associated with the preservation of beta-catenin levels. We asked whether mechanical up-regulation of beta-catenin was critical to reduction in adipogenesis as well as other mechanical events inducing alternate MSC lineage selection. In MSC cultured under strong adipogenic conditions, mechanical load (3600 cycles/day, 2% strain) inactivated GSK3beta in a Wnt-independent fashion. Small interfering RNA targeting GSK3beta prevented both strain-induced induction of beta-catenin and an increase in COX2, a factor associated with increased osteoprogenitor phenotype. Small interfering RNA knockdown of beta-catenin blocked mechanical reduction of peroxisome proliferator-activated receptor gamma and adiponectin, implicating beta-catenin in strain inhibition of adipogenesis. In contrast, the effect of both mechanical and pharmacologic inhibition of GSK3beta on the putative beta-catenin target, COX2, was unaffected by beta-catenin knockdown. GSK3beta inhibition caused accumulation of nuclear NFATc1; mechanical strain increased nuclear NFATc1, independent of beta-catenin. NFATc1 knockdown prevented mechanical stimulation of COX2, implicating NFATc1 signaling. Finally, inhibition of GSK3beta caused association of RNA polymerase II with the COX2 gene, suggesting transcription initiation. These results demonstrate that mechanical inhibition of GSK3beta induces activation of both beta-catenin and NFATc1 signaling, limiting adipogenesis via the former and promoting osteoblastic differentiation via NFATc1/COX2. Our novel findings suggest that mechanical loading regulates mesenchymal stem cell differentiation through inhibition of GSK3beta, which in turn regulates multiple downstream effectors.
Figures







Similar articles
-
Mechanical strain inhibits adipogenesis in mesenchymal stem cells by stimulating a durable beta-catenin signal.Endocrinology. 2008 Dec;149(12):6065-75. doi: 10.1210/en.2008-0687. Epub 2008 Aug 7. Endocrinology. 2008. PMID: 18687779 Free PMC article.
-
The effects of myostatin on adipogenic differentiation of human bone marrow-derived mesenchymal stem cells are mediated through cross-communication between Smad3 and Wnt/beta-catenin signaling pathways.J Biol Chem. 2008 Apr 4;283(14):9136-45. doi: 10.1074/jbc.M708968200. Epub 2008 Jan 18. J Biol Chem. 2008. PMID: 18203713 Free PMC article.
-
β-catenin directly sequesters adipocytic and insulin sensitizing activities but not osteoblastic activity of PPARγ2 in marrow mesenchymal stem cells.PLoS One. 2012;7(12):e51746. doi: 10.1371/journal.pone.0051746. Epub 2012 Dec 18. PLoS One. 2012. PMID: 23272157 Free PMC article.
-
PPARγ and Wnt Signaling in Adipogenic and Osteogenic Differentiation of Mesenchymal Stem Cells.Curr Stem Cell Res Ther. 2016;11(3):216-25. doi: 10.2174/1574888x10666150519093429. Curr Stem Cell Res Ther. 2016. PMID: 25986621 Review.
-
Molecular switching of osteoblastogenesis versus adipogenesis: implications for targeted therapies.Expert Opin Ther Targets. 2009 May;13(5):593-603. doi: 10.1517/14728220902915310. Expert Opin Ther Targets. 2009. PMID: 19397478 Review.
Cited by
-
P2X7 nucleotide receptor signaling potentiates the Wnt/β-catenin pathway in cells of the osteoblast lineage.Purinergic Signal. 2016 Sep;12(3):509-20. doi: 10.1007/s11302-016-9517-4. Epub 2016 May 21. Purinergic Signal. 2016. PMID: 27206526 Free PMC article.
-
Beta-catenin--a supporting role in the skeleton.J Cell Biochem. 2010 Jun 1;110(3):545-53. doi: 10.1002/jcb.22574. J Cell Biochem. 2010. PMID: 20512915 Free PMC article. Review.
-
Primary Cilia Exhibit Mechanosensitivity to Cyclic Tensile Strain and Lineage-Dependent Expression in Adipose-Derived Stem Cells.Sci Rep. 2019 May 29;9(1):8009. doi: 10.1038/s41598-019-43351-y. Sci Rep. 2019. PMID: 31142808 Free PMC article.
-
Chromatin accessibility and epigenetic deoxyribose nucleic acid (DNA) modifications in chronic kidney disease (CKD) osteoblasts: a study of bone and osteoblasts from pediatric patients with CKD.JBMR Plus. 2024 Jan 4;8(1):ziad015. doi: 10.1093/jbmrpl/ziad015. eCollection 2024 Jan. JBMR Plus. 2024. PMID: 38694428 Free PMC article.
-
Iron Oxide Nanoparticles Combined with Static Magnetic Fields in Bone Remodeling.Cells. 2022 Oct 20;11(20):3298. doi: 10.3390/cells11203298. Cells. 2022. PMID: 36291164 Free PMC article. Review.
References
-
- Minaire P., Edouard C., Arlot M., Meunier P. J. (1984) Calcif. Tissue Int. 36, 338–340 - PubMed
-
- Shih T. T., Chang C. J., Hsu C. Y., Wei S. Y., Su K. C., Chung H. W. (2004) Spine 29, 2844–2850 - PubMed
-
- David V., Martin A., Lafage-Proust M. H., Malaval L., Peyroche S., Jones D. B., Vico L., Guignandon A. (2007) Endocrinology 148, 2553–2562 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
