

TFIIF facilitates dissociation of RNA polymerase II from noncoding RNAs that lack a repression domain
- PMID: 19841064
- PMCID: PMC2798307
- DOI: 10.1128/MCB.01115-09
TFIIF facilitates dissociation of RNA polymerase II from noncoding RNAs that lack a repression domain
Abstract
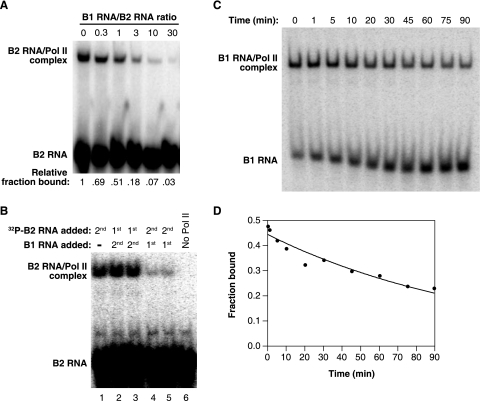
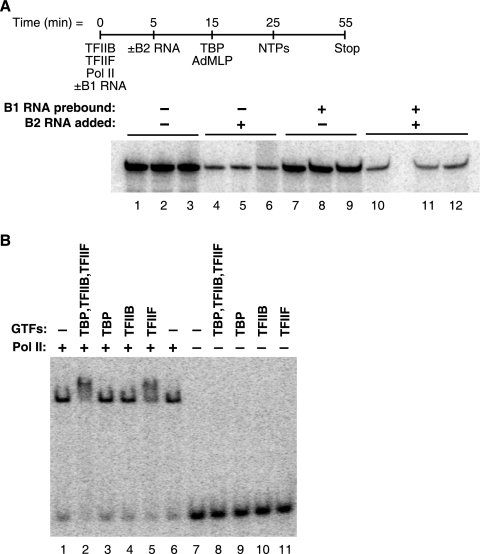
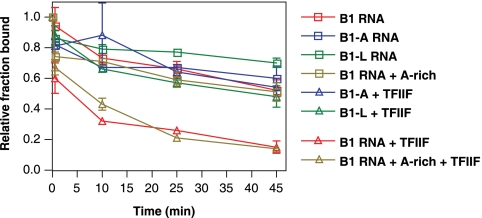
Noncoding RNAs (ncRNAs) have recently been found to regulate multiple steps in mammalian mRNA transcription. Mouse B2 RNA and human Alu RNA bind RNA polymerase II (Pol II) and repress mRNA transcription, using regions of the ncRNAs referred to as repression domains. Two other ncRNAs, mouse B1 RNA and human small cytoplasmic Alu (scAlu) RNA, bind Pol II with high affinity but lack repression domains and hence do not inhibit transcription. To better understand the interplay between ncRNAs that bind Pol II and their functions in transcription, we studied how Pol II binding and transcriptional repression are controlled by general transcription factors. We found that TFIIF associates with B1 RNA/Pol II and scAlu RNA/Pol II complexes and decreases their kinetic stability. Both subunits of TFIIF are required for this activity. Importantly, fusing a repression domain to B1 RNA stabilizes its interaction with Pol II in the presence of TFIIF. These results suggest a new role for TFIIF in regulating the interaction of ncRNAs with Pol II; specifically, it destabilizes interactions with ncRNAs that are not transcriptional repressors. These studies also identify a new function for ncRNA repression domains: they stabilize interactions of ncRNAs with Pol II in the presence of TFIIF.
Figures



References
-
- Allen, T. A., S. Von Kaenel, J. A. Goodrich, and J. F. Kugel. 2004. The SINE-encoded mouse B2 RNA represses mRNA transcription in response to heat shock. Nat. Struct. Mol. Biol. 11:816-821. - PubMed
-
- Barrandon, C., B. Spiluttini, and O. Bensaude. 2008. Non-coding RNAs regulating the transcriptional machinery. Biol. Cell 100:83-95. - PubMed
-
- Chang, C.-H., C. F. Kostrub, and Z. F. Burton. 1993. RAP30/74 (transcription factor IIF) is required for promoter escape by RNA polymerase II. J. Biol. Chem. 268:20482-20489. - PubMed
-
- Conaway, J. W., and R. C. Conaway. 1990. An RNA polymerase II transcription factor shares functional properties with Escherichia coli sigma 70. Science 248:1550-1553. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases