

Cytochrome P450 125 (CYP125) catalyses C26-hydroxylation to initiate sterol side-chain degradation in Rhodococcus jostii RHA1
- PMID: 19843222
- PMCID: PMC5218833
- DOI: 10.1111/j.1365-2958.2009.06915.x
Cytochrome P450 125 (CYP125) catalyses C26-hydroxylation to initiate sterol side-chain degradation in Rhodococcus jostii RHA1
Abstract
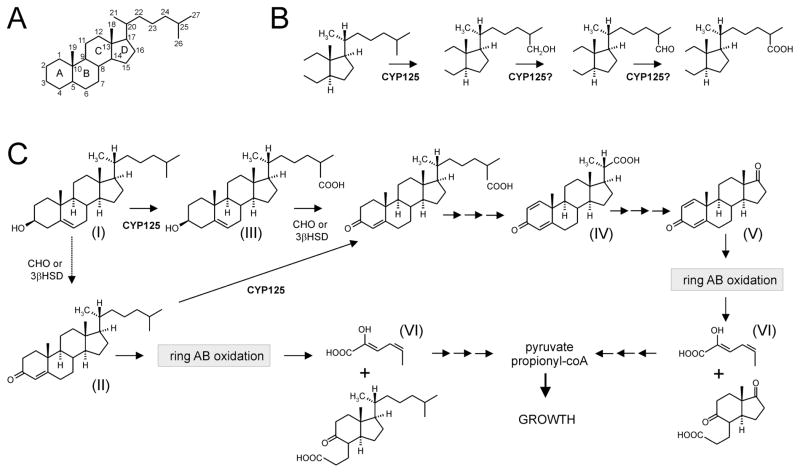
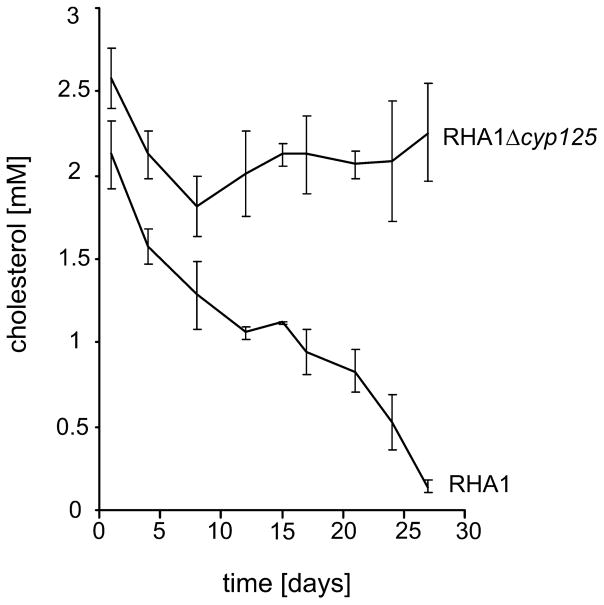
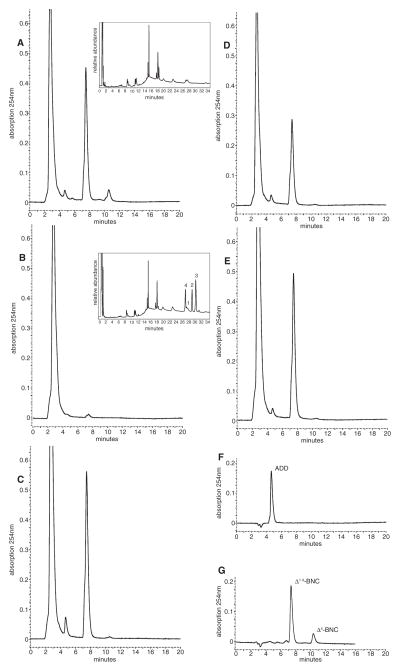
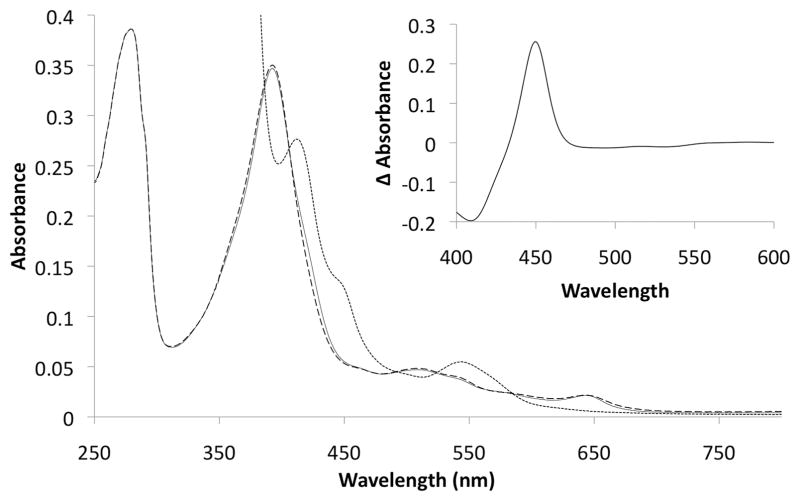
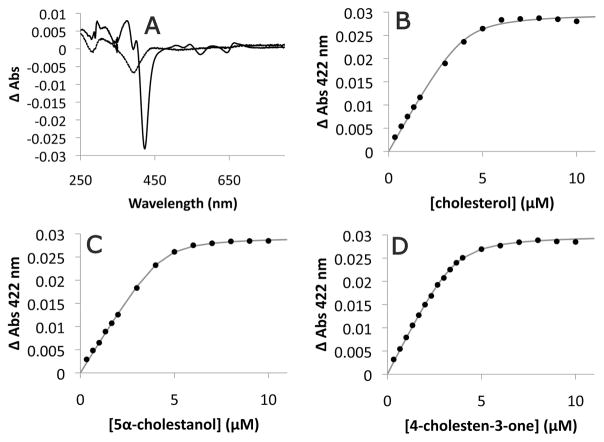
The cyp125 gene of Rhodococcus jostii RHA1 was previously found to be highly upregulated during growth on cholesterol and the orthologue in Mycobacterium tuberculosis (rv3545c) has been implicated in pathogenesis. Here we show that cyp125 is essential for R. jostii RHA1 to grow on 3-hydroxysterols such as cholesterol, but not on 3-oxo sterol derivatives, and that CYP125 performs an obligate first step in cholesterol degradation. The involvement of cyp125 in sterol side-chain degradation was confirmed by disrupting the homologous gene in Rhodococcus rhodochrous RG32, a strain that selectively degrades the cholesterol side-chain. The RG32 Omega cyp125 mutant failed to transform the side-chain of cholesterol, but degraded that of 5-cholestene-26-oic acid-3beta-ol, a cholesterol catabolite. Spectral analysis revealed that while purified ferric CYP125(RHA1) was < 10% in the low-spin state, cholesterol (K(D)(app) = 0.20 +/- 0.08 microM), 5 alpha-cholestanol (K(D)(app) = 0.15 +/- 0.03 microM) and 4-cholestene-3-one (K(D)(app) = 0.20 +/- 0.03 microM) further reduced the low spin character of the haem iron consistent with substrate binding. Our data indicate that CYP125 is involved in steroid C26-carboxylic acid formation, catalysing the oxidation of C26 either to the corresponding carboxylic acid or to an intermediate state.
Figures
References
-
- Ambrus G, Jekkel A, Ilkoy E, Horvath G, Bocskei Z. Novel 26-oxygenated products in microbial degradation of ergosterol. Steroids. 1995;60:626–629. - PubMed
-
- Arima K, Nakamatsu T, Beppu T. Microbial production of 3-oxobisnorchola-1,4-dien-22-oic acid. Agric Biol Chem. 1978;42:411–416.
-
- Camus JC, Pryor MJ, Medigue C, Cole ST. Re-annotation of the genome sequence of Mycobacterium tuberculosis H37Rv. Microbiology. 2002;148:2967–2973. - PubMed
-
- Chang JC, Harik NS, Liao RP, Sherman DR. Identification of Mycobacterial genes that alter growth and pathology in macrophages and in mice. J Infect Dis. 2007;196:788–795. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
