

Nuclear phosphoinositides: a signaling enigma wrapped in a compartmental conundrum
- PMID: 19846310
- PMCID: PMC2818233
- DOI: 10.1016/j.tcb.2009.09.009
Nuclear phosphoinositides: a signaling enigma wrapped in a compartmental conundrum
Abstract
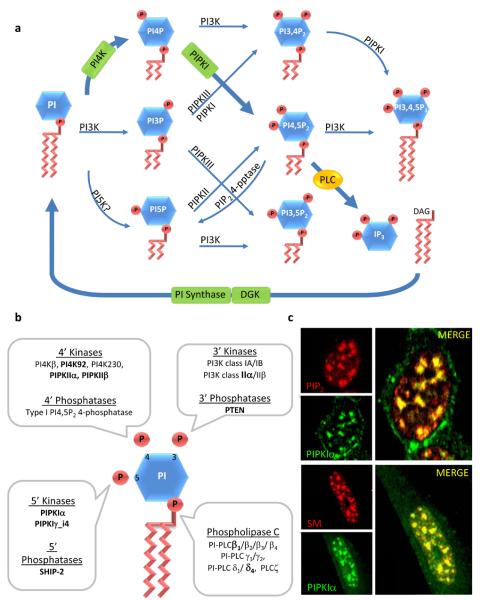
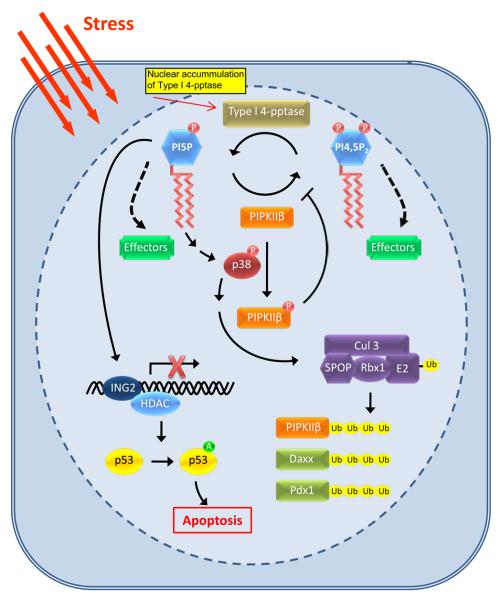
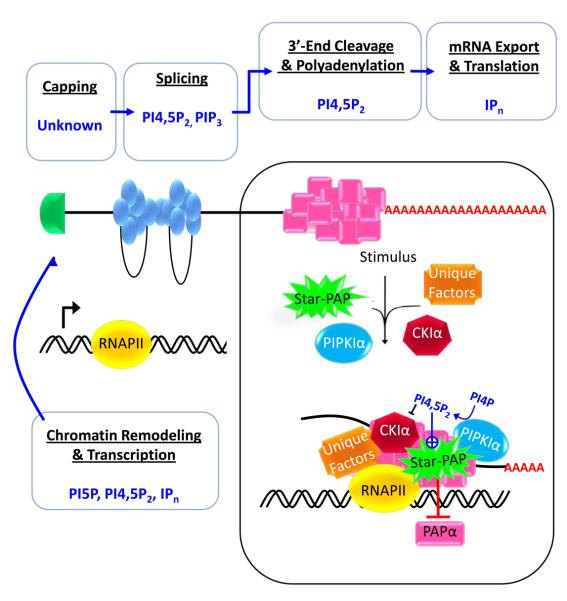
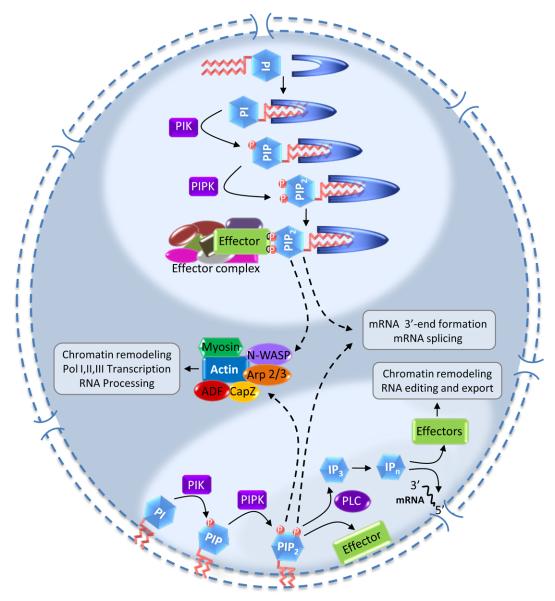
While the presence of phosphoinositides in the nuclei of eukaryotes and the identity of the enzymes responsible for their metabolism have been known for some time, their functions in the nucleus are only now emerging. This is illustrated by the recent identification of effectors for nuclear phosphoinositides. Like the cytosolic phosphoinositide signaling pathway, nuclear phosphatidylinositol 4,5-bisphosphate (PI4,5P(2)) is at the center of the pathway and acts both as a messenger and as a precursor for many additional messengers. Here, recent advances in the understanding of nuclear phosphoinositide signaling and its functions are reviewed with an emphasis on PI4,5P(2) and its role in gene expression. The compartmentalization of nuclear phosphoinositide phosphates (PIP(n)) remains a mystery, but emerging evidence suggests that phosphoinositides occupy several functionally distinct compartments.
Figures
References
-
- Hokin MR, Hokin LE. Enzyme secretion and the incorporation of P32 into phospholipides of pancreas slices. J Biol Chem. 1953;203:967–977. - PubMed
-
- Heck JN, et al. A conspicuous connection: structure defines function for the phosphatidylinositol-phosphate kinase family. Crit Rev Biochem Mol Biol. 2007;42:15–39. - PubMed
-
- Divecha N, et al. The polyphosphoinositide cycle exists in the nuclei of Swiss 3T3 cells under the control of a receptor (for IGF-I) in the plasma membrane, and stimulation of the cycle increases nuclear diacylglycerol and apparently induces translocation of protein kinase C to the nucleus. Embo J. 1991;10:3207–3214. - PMC - PubMed
-
- Irvine RF. Nuclear lipid signaling. Sci STKE. 2002;2002:RE13. - PubMed
-
- Irvine RF. Nuclear lipid signalling. Nat Rev Mol Cell Biol. 2003;4:349–360. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
