

T-type calcium channel inhibition underlies the analgesic effects of the endogenous lipoamino acids
- PMID: 19846698
- PMCID: PMC6665211
- DOI: 10.1523/JNEUROSCI.2919-09.2009
T-type calcium channel inhibition underlies the analgesic effects of the endogenous lipoamino acids
Abstract
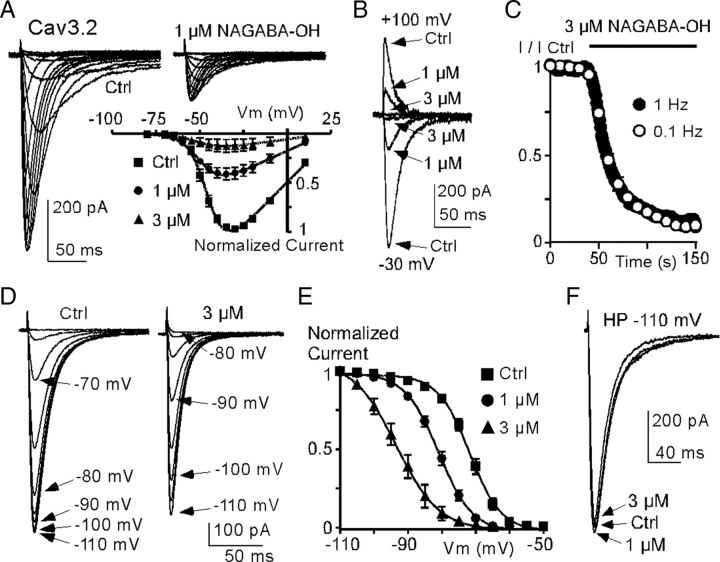
Lipoamino acids are anandamide-related endogenous molecules that induce analgesia via unresolved mechanisms. Here, we provide evidence that the T-type/Cav3 calcium channels are important pharmacological targets underlying their physiological effects. Various lipoamino acids, including N-arachidonoyl glycine (NAGly), reversibly inhibited Cav3.1, Cav3.2, and Cav3.3 currents, with potent effects on Cav3.2 [EC(50) approximately 200 nm for N-arachidonoyl 3-OH-gamma-aminobutyric acid (NAGABA-OH)]. This inhibition involved a large shift in the Cav3.2 steady-state inactivation and persisted during fatty acid amide hydrolase (FAAH) inhibition as well as in cell-free outside-out patch. In contrast, lipoamino acids had weak effects on high-voltage-activated (HVA) Cav1.2 and Cav2.2 calcium currents, on Nav1.7 and Nav1.8 sodium currents, and on anandamide-sensitive TRPV1 and TASK1 currents. Accordingly, lipoamino acids strongly inhibited native Cav3.2 currents in sensory neurons with small effects on sodium and HVA calcium currents. In addition, we demonstrate here that lipoamino acids NAGly and NAGABA-OH produced a strong thermal analgesia and that these effects (but not those of morphine) were abolished in Cav3.2 knock-out mice. Collectively, our data revealed lipoamino acids as a family of endogenous T-type channel inhibitors, suggesting that these ligands can modulate multiple cell functions via this newly evidenced regulation.
Figures




References
-
- Beedle AM, McRory JE, Poirot O, Doering CJ, Altier C, Barrere C, Hamid J, Nargeot J, Bourinet E, Zamponi GW. Agonist-independent modulation of N-type calcium channels by ORL1 receptors. Nat Neurosci. 2004;7:118–125. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources