

Role of TRPC1 channel in skeletal muscle function
- PMID: 19846750
- PMCID: PMC2806157
- DOI: 10.1152/ajpcell.00241.2009
Role of TRPC1 channel in skeletal muscle function
Abstract
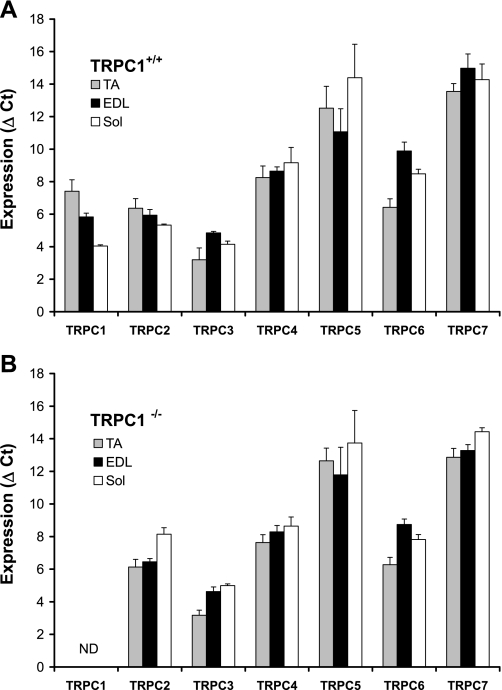
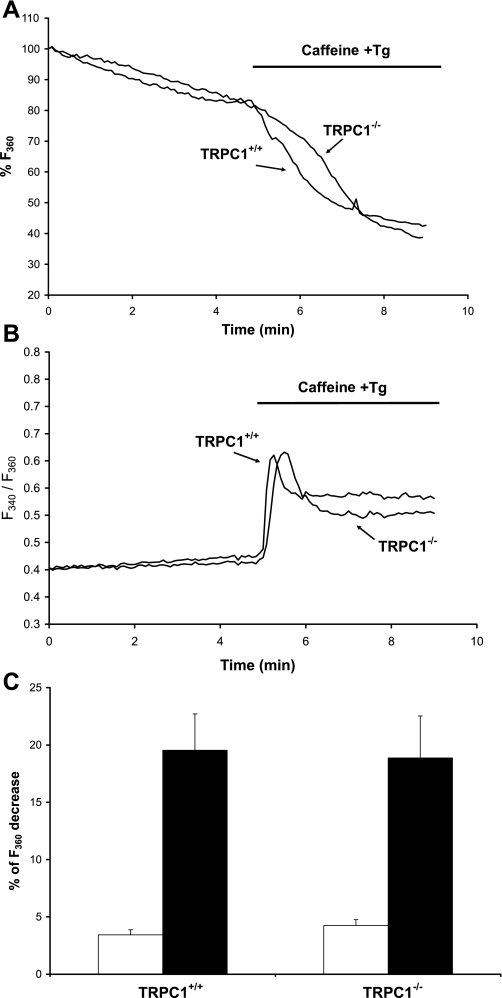
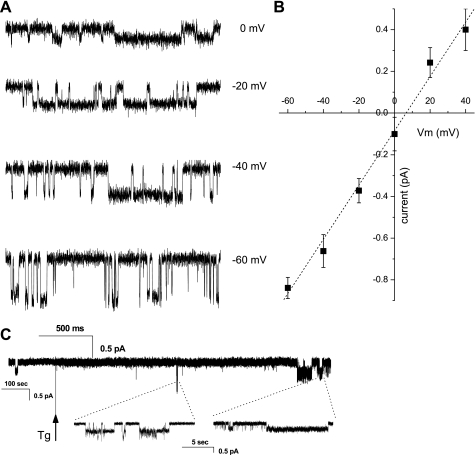
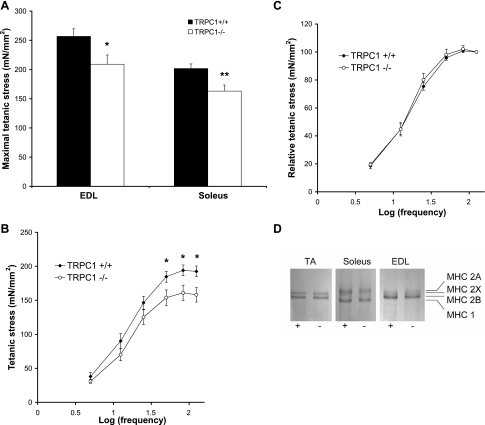
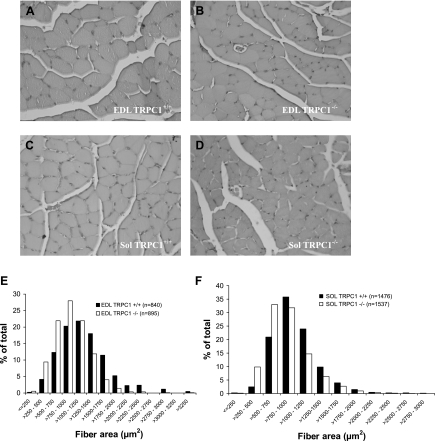
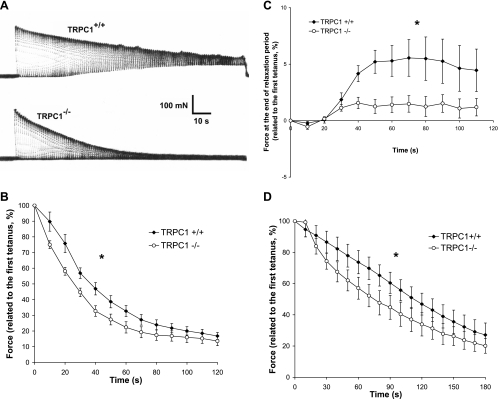
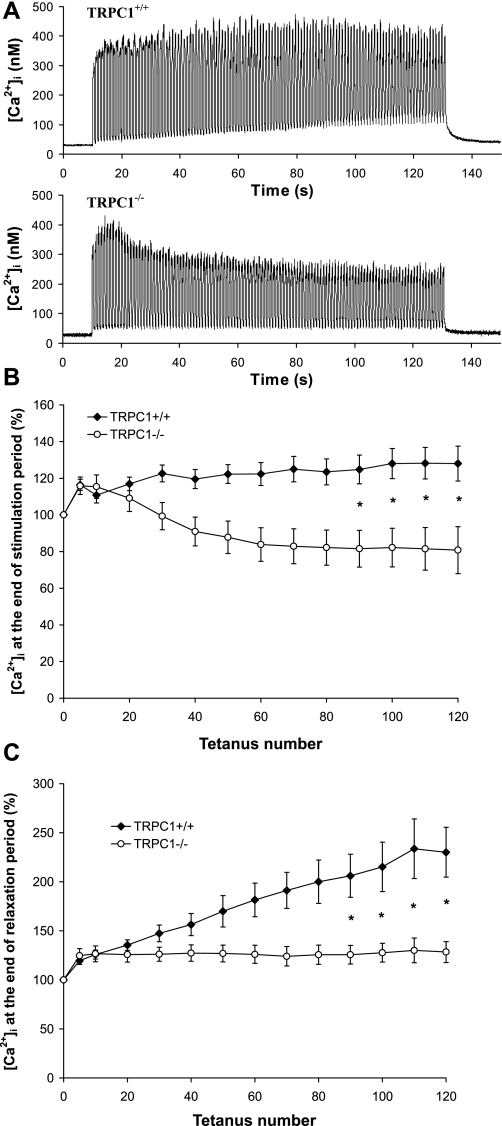
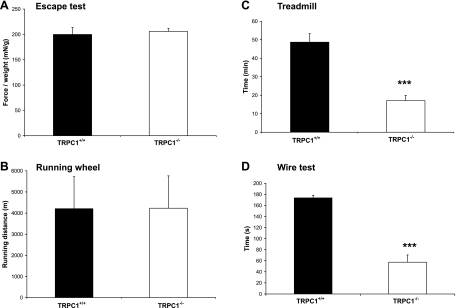
Skeletal muscle contraction is reputed not to depend on extracellular Ca2+. Indeed, stricto sensu, excitation-contraction coupling does not necessitate entry of Ca2+. However, we previously observed that, during sustained activity (repeated contractions), entry of Ca2+ is needed to maintain force production. In the present study, we evaluated the possible involvement of the canonical transient receptor potential (TRPC)1 ion channel in this entry of Ca2+ and investigated its possible role in muscle function. Patch-clamp experiments reveal the presence of a small-conductance channel (13 pS) that is completely lost in adult fibers from TRPC1(-/-) mice. The influx of Ca2+ through TRPC1 channels represents a minor part of the entry of Ca(2+) into muscle fibers at rest, and the activity of the channel is not store dependent. The lack of TRPC1 does not affect intracellular Ca2+ concentration ([Ca2+](i)) transients reached during a single isometric contraction. However, the involvement of TRPC1-related Ca2+ entry is clearly emphasized in muscle fatigue. Indeed, muscles from TRPC1(-/-) mice stimulated repeatedly progressively display lower [Ca2+](i) transients than those observed in TRPC1(+/+) fibers, and they also present an accentuated progressive loss of force. Interestingly, muscles from TRPC1(-/-) mice display a smaller fiber cross-sectional area, generate less force per cross-sectional area, and contain less myofibrillar proteins than their controls. They do not present other signs of myopathy. In agreement with in vitro experiments, TRPC1(-/-) mice present an important decrease of endurance of physical activity. We conclude that TRPC1 ion channels modulate the entry of Ca(2+) during repeated contractions and help muscles to maintain their force during sustained repeated contractions.
Figures
References
-
- Allen DG. Skeletal muscle function: role of ionic changes in fatigue, damage and disease. Clin Exp Pharmacol Physiol 31: 485–493, 2004 - PubMed
-
- Allen DG, Lamb GD, Westerblad H. Skeletal muscle fatigue: cellular mechanisms. Physiol Rev 88: 287–332, 2008 - PubMed
-
- Allen DG, Lannergren J, Westerblad H. Muscle cell function during prolonged activity: cellular mechanisms of fatigue. Exp Physiol 80: 497–527, 1995 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous