

Modulation of genioglossus muscle activity across sleep-wake states by histamine at the hypoglossal motor pool
- PMID: 19848360
- PMCID: PMC2753809
- DOI: 10.1093/sleep/32.10.1313
Modulation of genioglossus muscle activity across sleep-wake states by histamine at the hypoglossal motor pool
Abstract
Study objectives: Histamine neurons comprise a major component of the aminergic arousal system and significantly influence sleep-wake states, with antihistamines widely used as sedative hypnotics. Unlike the serotonergic and noradrenergic components of this arousal system, however, the role of histamine in the central control of respiratory motor activity has not been determined. The aims of this study were to characterize the effects of histamine receptor agonists and antagonists at the hypoglossal motor pool on genioglossus muscle activity across sleep and awake states, and also determine if histamine contributes an endogenous excitatory drive to modulate hypoglossal motor outflow to genioglossus muscle.
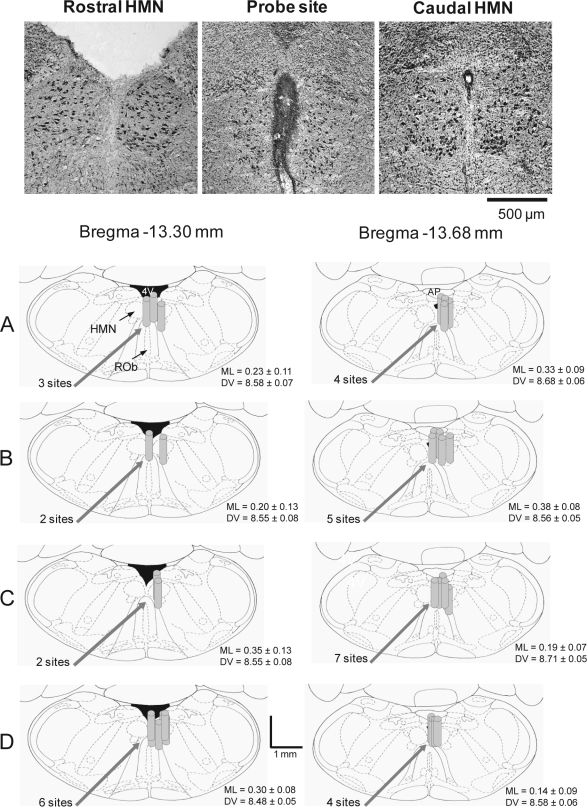
Design, participants, and interventions: Thirty-three rats were implanted with electroencephalogram and neck electrodes to record sleep-wake states, and genioglossus and diaphragm electrodes for respiratory muscle recordings. Microdialysis probes were inserted into the hypoglossal motor nucleus.
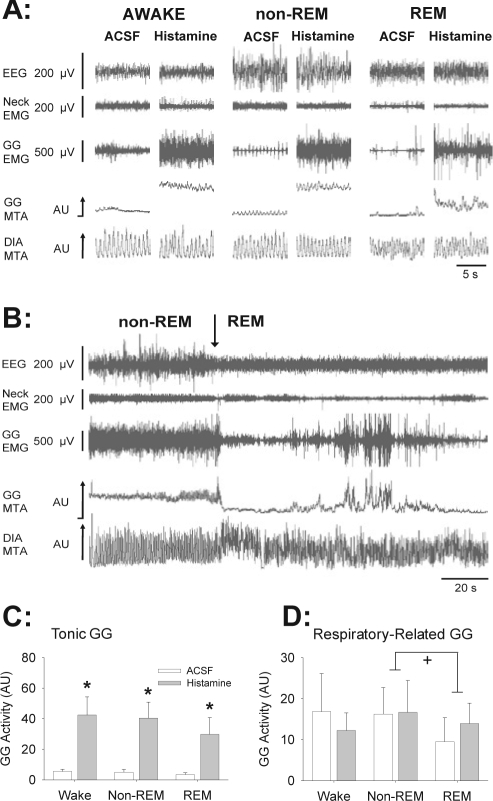
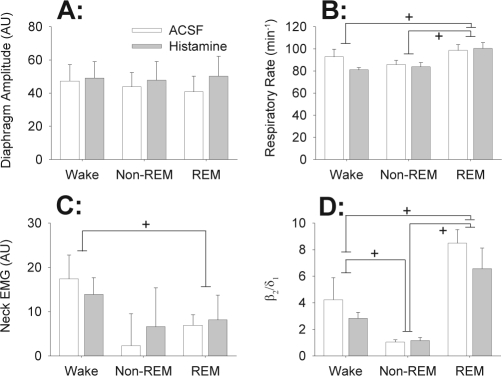
Measurements and results: Histamine at the hypoglossal motor nucleus significantly increased tonic genioglossus muscle activity in wakefulness, non-REM sleep and REM sleep. The activating effects of histamine on genioglossus muscle activity also occurred with a histamine type-1 (H1) but not H2 receptor agonist. However, H1 receptor antagonism at the hypoglossal motor nucleus did not decrease genioglossus muscle activity in wakefulness or sleep.
Conclusions: The results suggest that histamine at the hypoglossal motor pool increases genioglossus muscle activity in freely behaving rats in wakefulness, non-REM, and REM sleep via an H1 receptor mechanism.
Figures



References
-
- Jones BE. Basic mechanisms of sleep-wake states. In: Kryger MH, Roth T, Dement WC, editors. Principles and practice of sleep medicine. 2nd ed. Philadelphia: Saunders; 1994. pp. 145–62.
-
- Jacobs BL, Azmitia EC. Structure and function of the brain serotonin system. Physiol Rev. 1992;72:165–229. - PubMed
-
- Foote SL, Bloom FE, Aston-Jones G. Nucleus locus ceruleus: new evidence of anatomical and physiological specificity. Physiol Rev. 1983;63:844–914. - PubMed
-
- Saper CB, Scammell TE, Lu J. Hypothalamic regulation of sleep and circadian rhythms. Nature. 2005;437:1257–63. - PubMed
