

An activity-induced microRNA controls dendritic spine formation by regulating Rac1-PAK signaling
- PMID: 19850129
- PMCID: PMC2818337
- DOI: 10.1016/j.mcn.2009.10.005
An activity-induced microRNA controls dendritic spine formation by regulating Rac1-PAK signaling
Erratum in
- Mol Cell Neurosci. 2012 Feb;49(2):250. Lasiek, Adam [corrected to Lesiak, Adam]
Abstract
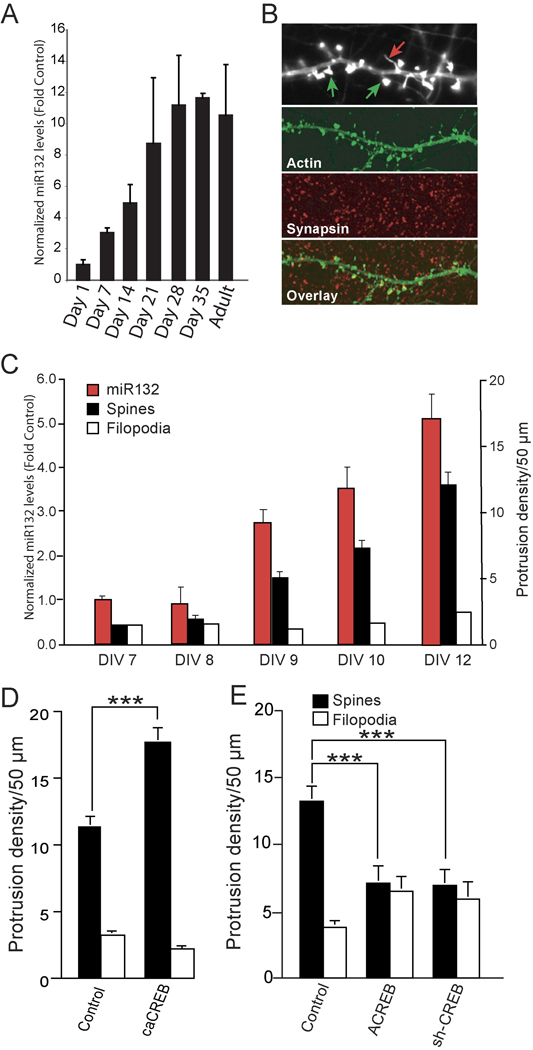
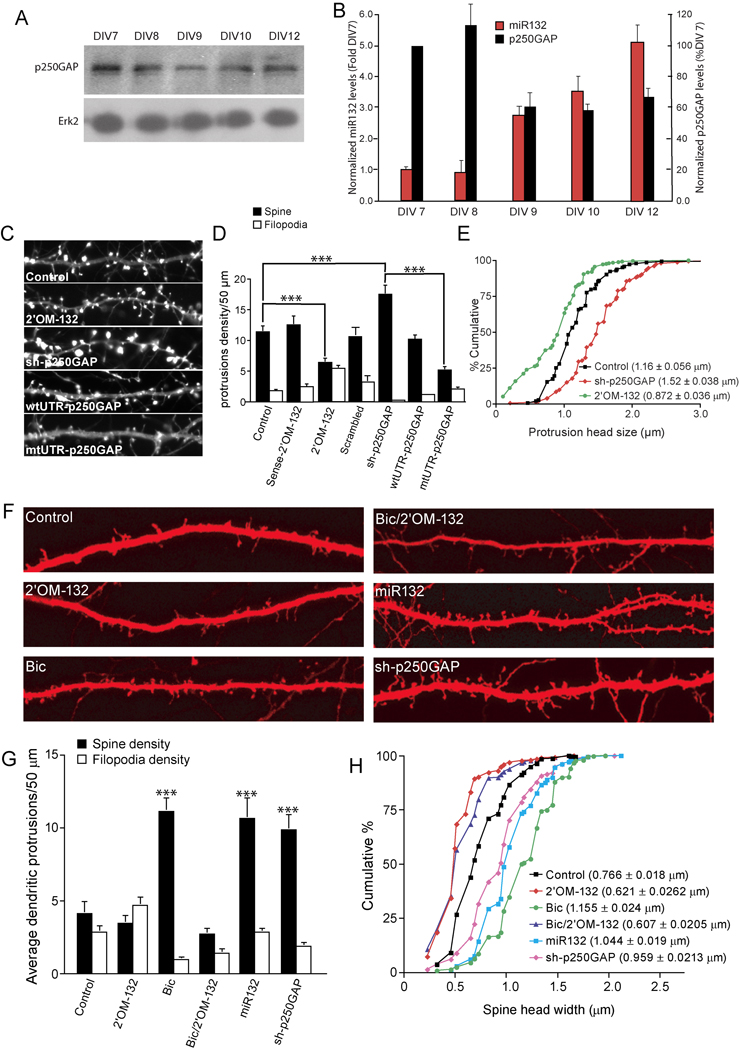
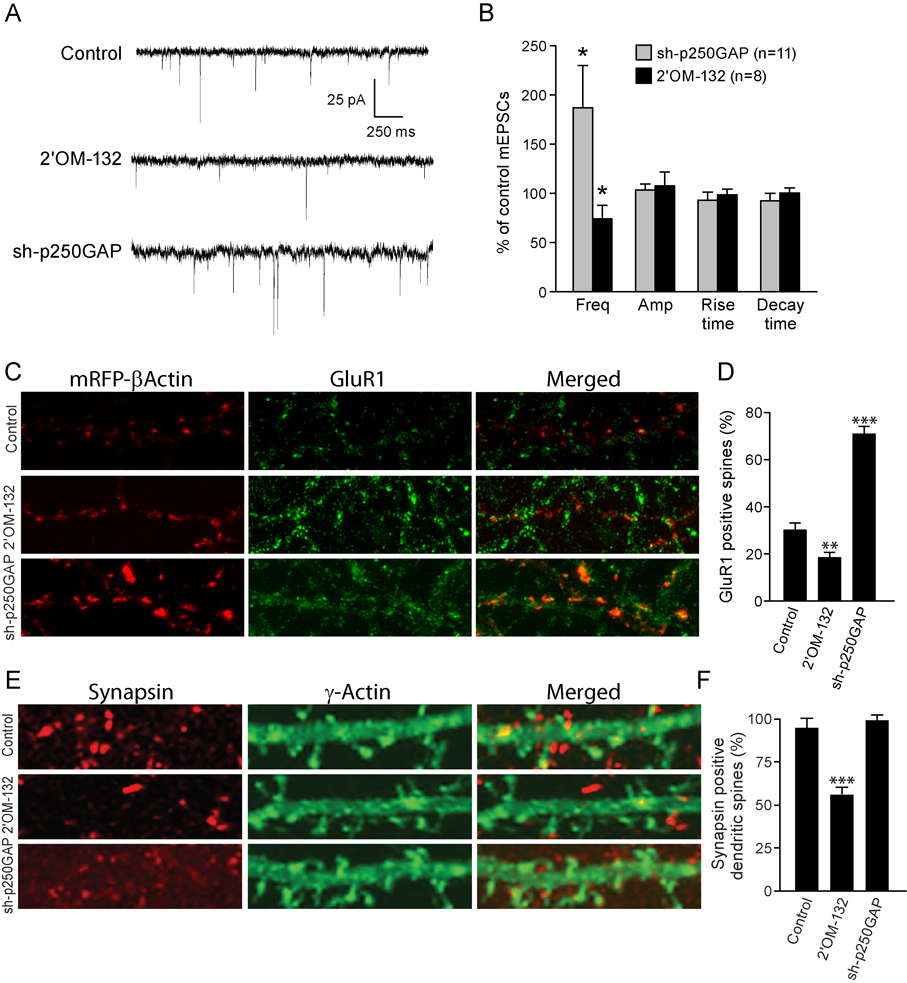
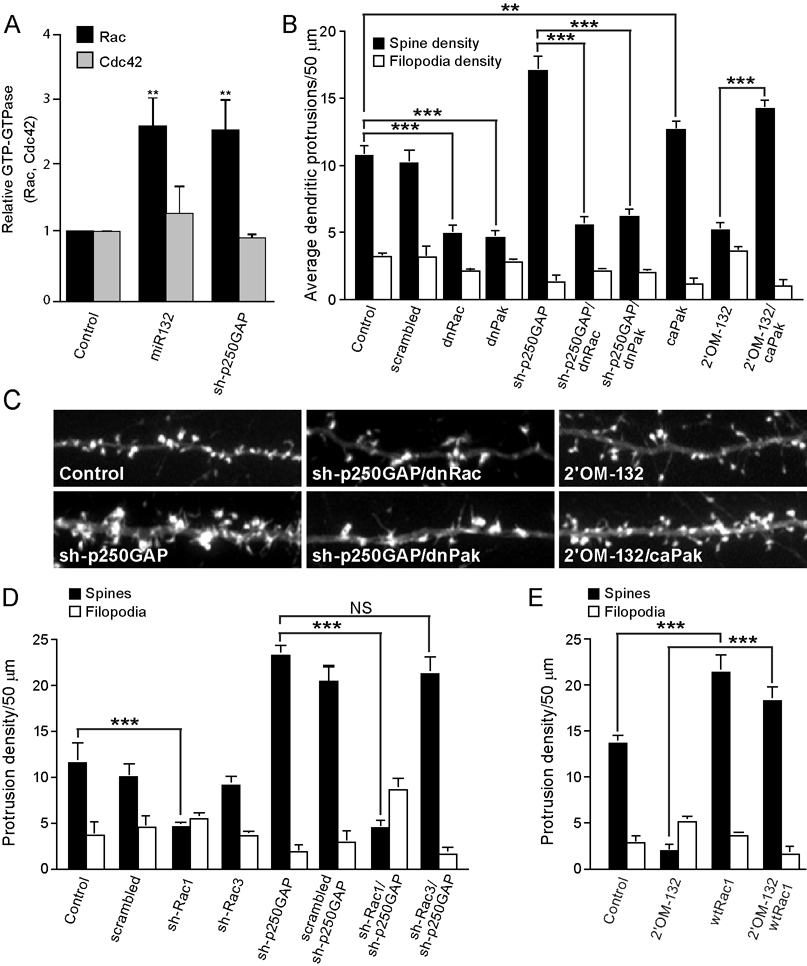
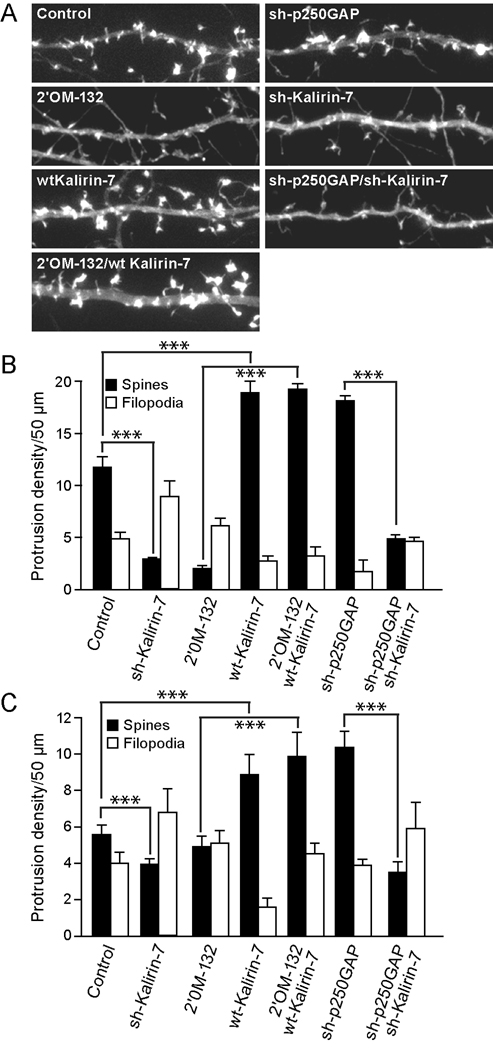
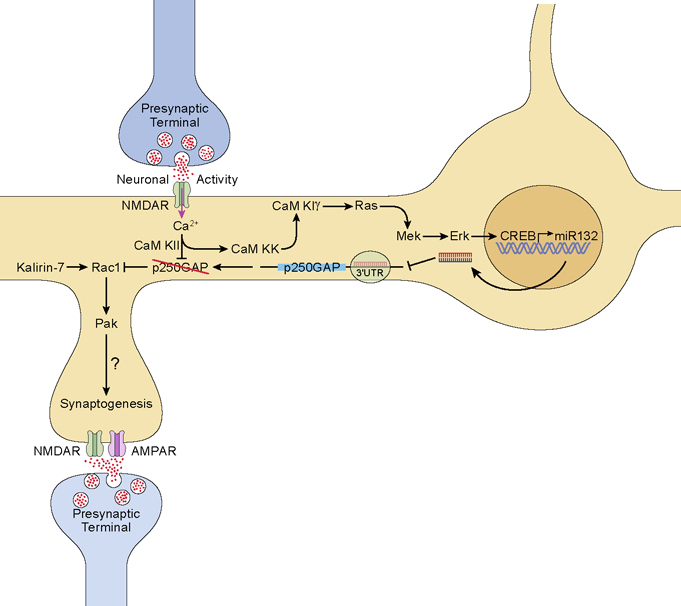
Activity-regulated gene expression is believed to play a key role in the development and refinement of neuronal circuitry. Nevertheless, the transcriptional networks that regulate synaptic plasticity remain largely uncharacterized. We show here that the CREB- and activity-regulated microRNA, miR132, is induced during periods of active synaptogenesis. Moreover, miR132 is necessary and sufficient for hippocampal spine formation. Expression of the miR132 target, p250GAP, is inversely correlated with miR132 levels and spinogenesis. Furthermore, knockdown of p250GAP increases spine formation while introduction of a p250GAP mutant unresponsive to miR132 attenuates this activity. Inhibition of miR132 decreases both mEPSC frequency and the number of GluR1-positive spines, while knockdown of p250GAP has the opposite effect. Additionally, we show that the miR132/p250GAP circuit regulates Rac1 activity and spine formation by modulating synapse-specific Kalirin7-Rac1 signaling. These data suggest that neuronal activity regulates spine formation, in part, by increasing miR132 transcription, which in turn activates a Rac1-Pak actin remodeling pathway.
Published by Elsevier Inc.
Figures
References
-
- Armstrong D, Dunn JK, Antalffy B, Trivedi R. Selective dendritic alterations in the cortex of Rett syndrome. J Neuropathol Exp Neurol. 1995;54:195–201. - PubMed
-
- Ashraf SI, McLoon AL, Sclarsic SM, Kunes S. Synaptic protein synthesis associated with memory is regulated by the RISC pathway in Drosophila. Cell. 2006;124:191–205. - PubMed
-
- Bahr BA. Long-term hippocampal slices: a model system for investigating synaptic mechanisms and pathologic processes. J Neurosci Res. 1995;42:294–305. - PubMed
-
- Bartsch D, Casadio A, Karl KA, Serodio P, Kandel ER. CREB1 encodes a nuclear activator, a repressor, and a cytoplasmic modulator that form a regulatory unit critical for long-term facilitation. Cell. 1998;95:211–223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
