

Primary microcephaly: do all roads lead to Rome?
- PMID: 19850369
- PMCID: PMC2816178
- DOI: 10.1016/j.tig.2009.09.011
Primary microcephaly: do all roads lead to Rome?
Abstract
The relatively large brain and expanded cerebral cortex of humans is unusual in the animal kingdom and is thought to have promoted our adaptability and success as a species. One approach for investigating neurogenesis is the study of autosomal recessive primary microcephaly (MCPH), in which prenatal brain growth is significantly reduced without an effect on brain structure. To date, eight MCPH loci and five genes have been identified. Unexpectedly, all MCPH proteins are ubiquitous and localise to centrosomes for at least part of the cell cycle. Here, we focus on recent functional studies of MCPH proteins that reveal the centrosome as a final integration point for many regulatory pathways affecting prenatal neurogenesis in mammals.
Figures
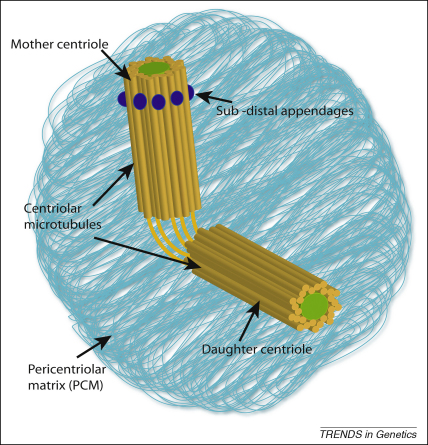
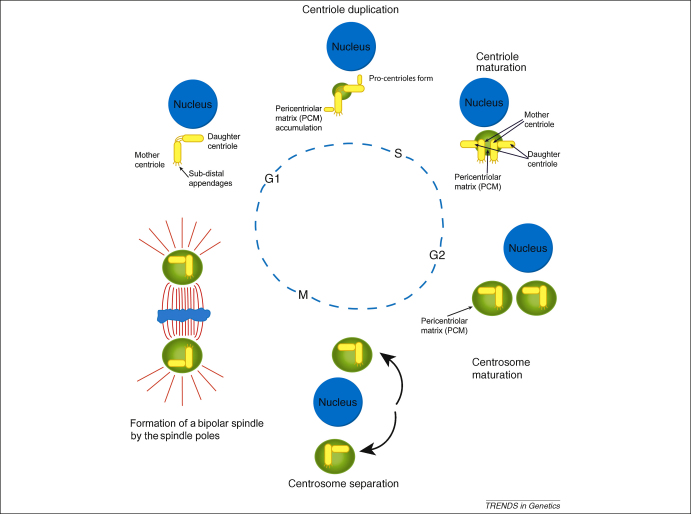
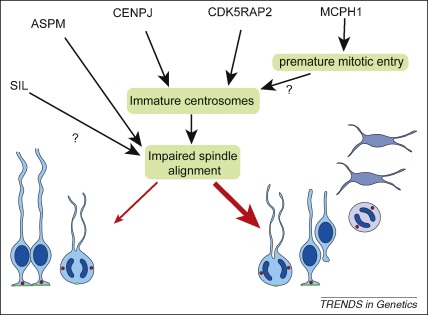
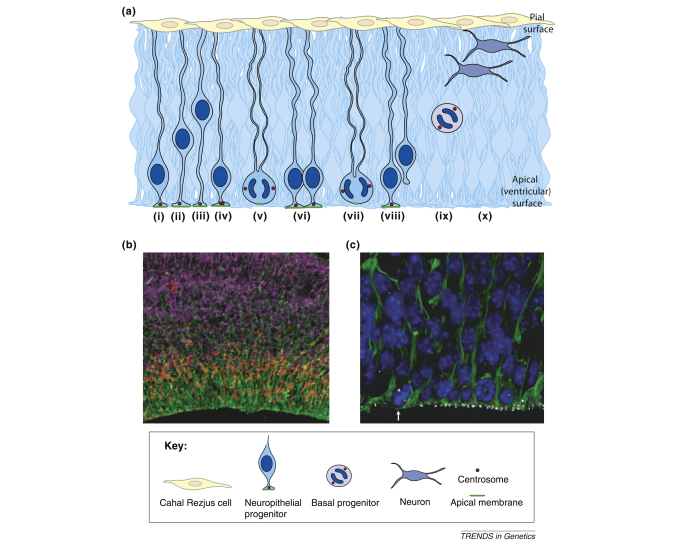
References
-
- Cox J. What primary microcephaly can tell us about brain growth. Trends Mol. Med. 2006;12:358–366. - PubMed
-
- Barkovich J.A. A developmental classification of malformations of the brainstem. Ann. Neurol. 1997;62:625–639. - PubMed
-
- Desir J. Primary microcephaly with ASPM mutation shows simplified cortical gyration with antero-posterior gradient pre- and post-natally. Am. J. Med. Genet. A. 2008;146A:1439–1443. - PubMed
-
- Saadi A. Compound heterozygous ASPM mutations associated with microcephaly and simplified cortical gyration in a consanguineous Algerian family. Eur. J. Med. Genet. 2008;52:180–184. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
