

Small ubiquitin-like modifier-2 modification of retinoic acid receptor-alpha regulates its subcellular localization and transcriptional activity
- PMID: 19850744
- PMCID: PMC2795702
- DOI: 10.1210/en.2009-0868
Small ubiquitin-like modifier-2 modification of retinoic acid receptor-alpha regulates its subcellular localization and transcriptional activity
Abstract
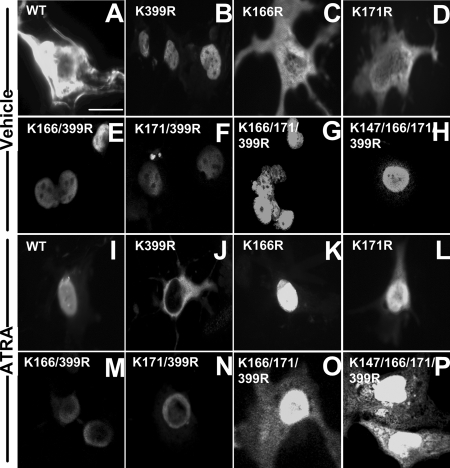
The retinoic acid receptor-alpha (Rara) gene is critical for germ cell development in the testis, as demonstrated by infertile Rara knockout male mice. The encoded protein for Rara (RARA) is expressed in both Sertoli cells and germ cells, but it is not always in the nucleus. Previously, all-trans retinoic acid (ATRA) was shown to increase the nuclear localization and transcriptional activity of RARA in Sertoli cells. Here, we identified a small ubiquitin-like modifier-2 (SUMO-2) modification as a novel posttranslational regulatory mechanism controlling the ATRA-dependent RARA subcellular localization and transcription. ATRA increased the SUMO-2 modification of RARA. In the presence of ATRA, lysine 166 (K166) and K171 of RARA were modified at a physiological concentration of SUMO-2, whereas in the absence of ATRA, K399 was the only site that was modified, but at a higher SUMO-2 concentration. However, K399 was critical for ATRA-controlled nuclear trafficking of RARA. In the presence of ATRA, a K399 mutation to arginine resulted in the cytoplasmic localization of K399R mutant, indicating that K166 and K171 sumoylations were inhibitory to nuclear localization. This may be due to SUMO/sentrin-specific peptidase 6 (SENP6) not being able to bind K399R mutant to desumoylate K166 and K171 in Sertoli cells, whereas it can bind RARA with intact K399. On the other hand, functional K166 and K171 sites for sumoylation were required for a full transcriptional activity, when K399 was intact. These results together suggest that both K166 and K171 sumoylation and desumoylation are critical for optimal RARA function.
Figures





Similar articles
-
Retinoic acid modulates the subcellular localization of small ubiquitin-related modifier-2/3 (SUMO-2/3) in the testis.J Androl. 2010 Jul-Aug;31(4):406-18. doi: 10.2164/jandrol.109.008763. Epub 2010 Jan 14. J Androl. 2010. PMID: 20075418
-
Activity of retinoic acid receptor-alpha is directly regulated at its protein kinase A sites in response to follicle-stimulating hormone signaling.Endocrinology. 2010 May;151(5):2361-72. doi: 10.1210/en.2009-1338. Epub 2010 Mar 9. Endocrinology. 2010. PMID: 20215566 Free PMC article.
-
Disulfide isomerase glucose-regulated protein 58 is required for the nuclear localization and degradation of retinoic acid receptor alpha.Reproduction. 2010 Apr;139(4):717-31. doi: 10.1530/REP-09-0527. Epub 2010 Feb 3. Reproduction. 2010. PMID: 20130111
-
Novel treatment of acute promyelocytic leukemia: As₂O₃, retinoic acid and retinoid pharmacology.Curr Pharm Biotechnol. 2013;14(9):849-58. doi: 10.2174/1389201015666140113095812. Curr Pharm Biotechnol. 2013. PMID: 24433507 Review.
-
Protein modification by SUMO.Annu Rev Biochem. 2004;73:355-82. doi: 10.1146/annurev.biochem.73.011303.074118. Annu Rev Biochem. 2004. PMID: 15189146 Review.
Cited by
-
Retinoid acid receptor expression is helpful to distinguish between adenoma and well-differentiated carcinoma in the thyroid.Virchows Arch. 2013 Jun;462(6):619-32. doi: 10.1007/s00428-013-1419-z. Epub 2013 May 4. Virchows Arch. 2013. PMID: 23639973
-
SENP3 grants tight junction integrity and cytoskeleton architecture in mouse Sertoli cells.Oncotarget. 2017 Apr 7;8(35):58430-58442. doi: 10.18632/oncotarget.16915. eCollection 2017 Aug 29. Oncotarget. 2017. PMID: 28938568 Free PMC article.
-
Regulation of the nucleocytoplasmic trafficking of viral and cellular proteins by ubiquitin and small ubiquitin-related modifiers.Biol Cell. 2012 Mar;104(3):121-38. doi: 10.1111/boc.201100105. Epub 2011 Dec 28. Biol Cell. 2012. PMID: 22188262 Free PMC article. Review.
-
Retinoid receptor turnover mediated by sumoylation, ubiquitination and the valosin-containing protein is disrupted in glioblastoma.Sci Rep. 2019 Nov 7;9(1):16250. doi: 10.1038/s41598-019-52696-3. Sci Rep. 2019. PMID: 31700049 Free PMC article.
-
Sumoylation and regulation of cardiac gene expression.Circ Res. 2010 Jul 9;107(1):19-29. doi: 10.1161/CIRCRESAHA.110.220491. Circ Res. 2010. PMID: 20616338 Free PMC article. Review.
References
-
- Doyle TJ, Braun KW, McLean DJ, Wright RW, Griswold MD, Kim KH 2007 Potential functions of retinoic acid receptor A in Sertoli cells and germ cells during spermatogenesis. Ann NY Acad Sci 1120:114–130 - PubMed
-
- Akmal KM, Dufour JM, Kim KH 1997 Retinoic acid receptor α gene expression in the rat testis: potential role during the prophase of meiosis and in the transition from round to elongating spermatids. Biol Reprod 56:549–556 - PubMed
-
- Dufour JM, Kim KH 1999 Cellular and subcellular localization of six retinoid receptors in rat testis during postnatal development: identification of potential heterodimeric receptors. Biol Reprod 61:1300–1308 - PubMed
-
- Akmal KM, Dufour JM, Vo M, Higginson S, Kim KH 1998 Ligand-dependent regulation of retinoic acid receptor α in rat testis: in vivo response to depletion and repletion of vitamin A. Endocrinology 139:1239–1248 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
