

Discrete contributions of elastic fiber components to arterial development and mechanical compliance
- PMID: 19850904
- PMCID: PMC2797476
- DOI: 10.1161/ATVBAHA.109.193227
Discrete contributions of elastic fiber components to arterial development and mechanical compliance
Abstract
Objective: Even though elastin and fibrillin-1 are the major structural components of elastic fibers, mutations in elastin and fibrillin-1 lead to narrowing of large arteries in supravalvular aortic stenosis and dilation of the ascending aorta in Marfan syndrome, respectively. A genetic approach was therefore used here to distinguish the differential contributions of elastin and fibrillin-1 to arterial development and compliance.
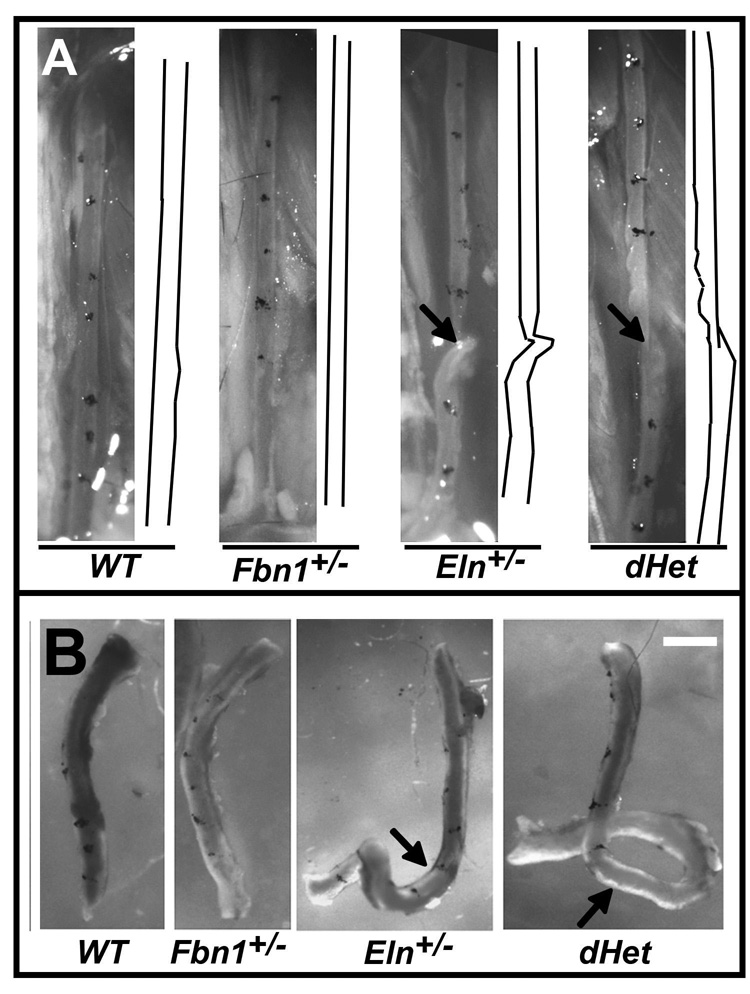
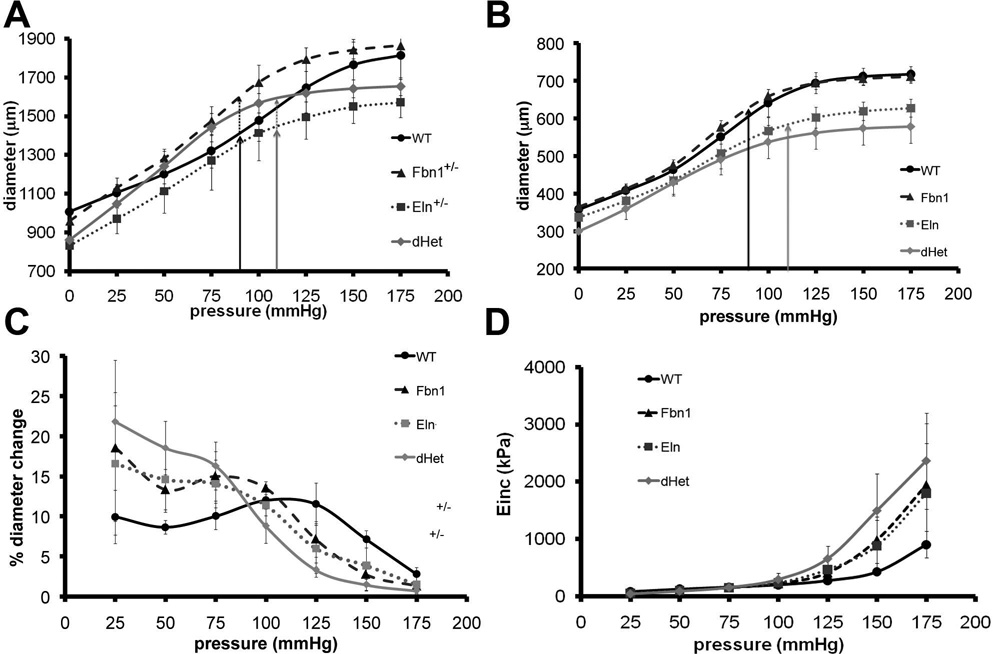
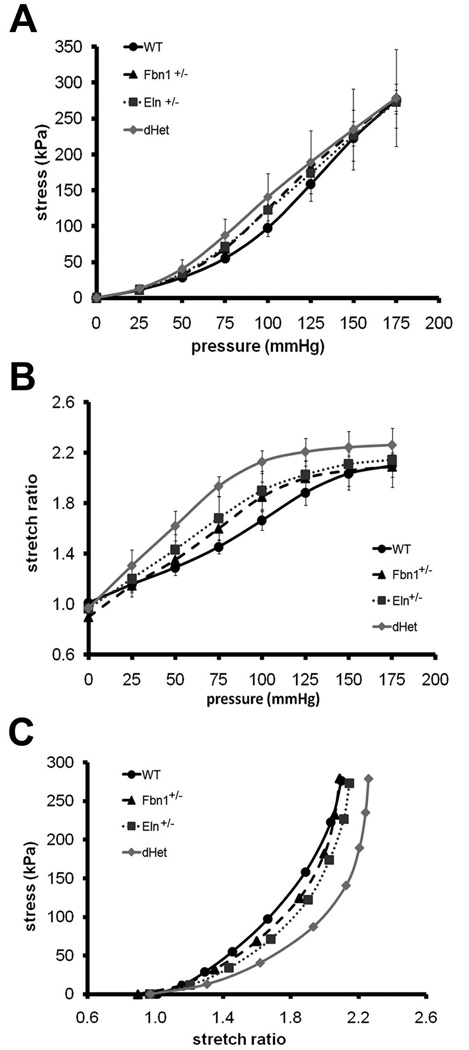
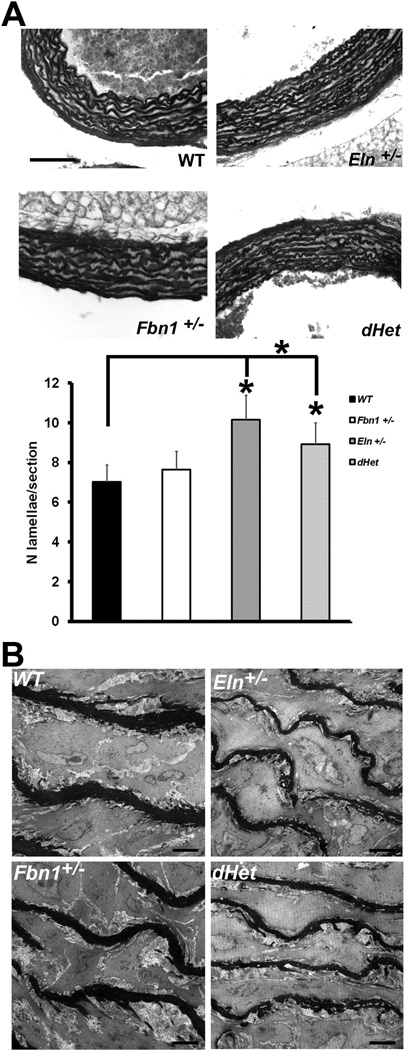
Methods and results: Key parameters of cardiovascular function were compared among adult mice haploinsufficient for elastin (Eln(+/-)), fibrillin-1 (Fbn1(+/-)), or both proteins (dHet). Physiological and morphological comparisons correlate elastin haploinsufficiency with increased blood pressure and vessel length and tortuosity in dHet mice, and fibrillin-1 haploinsufficiency with increased aortic diameter in the same mutant animals. Mechanical tests confirm that elastin and fibrillin-1 impart elastic recoil and tensile strength to the aortic wall, respectively. Additional ex vivo analyses demonstrate additive and overlapping contributions of elastin and fibrillin-1 to the material properties of vascular tissues. Lastly, light and electron microscopy evidence implicates fibrillin-1 in the hypertension-promoted remodeling of the elastin-deficient aorta.
Conclusions: These results demonstrate that elastin and fibrillin-1 have both differential and complementary roles in arterial wall formation and function, and advance our knowledge of the structural determinants of vascular physiology and disease.
Figures
Similar articles
-
Elastic and collagenous networks in vascular diseases.Cell Struct Funct. 2000 Apr;25(2):69-72. doi: 10.1247/csf.25.69. Cell Struct Funct. 2000. PMID: 10885576 Free PMC article. Review.
-
[Elastin and microfibrils in vascular development and ageing: complementary or opposite roles?].Biol Aujourdhui. 2012;206(2):87-102. doi: 10.1051/jbio/2012009. Epub 2012 Jul 4. Biol Aujourdhui. 2012. PMID: 22748047 Review. French.
-
Fibrillin-1 genetic deficiency leads to pathological ageing of arteries in mice.J Pathol. 2011 May;224(1):33-44. doi: 10.1002/path.2840. Epub 2011 Mar 22. J Pathol. 2011. PMID: 21432852 Free PMC article.
-
Basic components of connective tissues and extracellular matrix: elastin, fibrillin, fibulins, fibrinogen, fibronectin, laminin, tenascins and thrombospondins.Adv Exp Med Biol. 2014;802:31-47. doi: 10.1007/978-94-007-7893-1_3. Adv Exp Med Biol. 2014. PMID: 24443019 Review.
-
Mouse models of genetic diseases resulting from mutations in elastic fiber proteins.Matrix Biol. 2000 Nov;19(6):481-8. doi: 10.1016/s0945-053x(00)00101-3. Matrix Biol. 2000. PMID: 11068202 Review.
Cited by
-
Case report: Early carotid stent shortening in patient with radiation-induced carotid stenosis.Front Neurol. 2023 May 9;14:1184210. doi: 10.3389/fneur.2023.1184210. eCollection 2023. Front Neurol. 2023. PMID: 37228414 Free PMC article.
-
Decreased elastic energy storage, not increased material stiffness, characterizes central artery dysfunction in fibulin-5 deficiency independent of sex.J Biomech Eng. 2015 Mar;137(3):0310071-03100714. doi: 10.1115/1.4029431. Epub 2015 Jan 29. J Biomech Eng. 2015. PMID: 25532020 Free PMC article.
-
Minoxidil versus placebo in the treatment of arterial wall hypertrophy in children with Williams Beuren Syndrome: a randomized controlled trial.BMC Pediatr. 2019 May 28;19(1):170. doi: 10.1186/s12887-019-1544-1. BMC Pediatr. 2019. PMID: 31138170 Free PMC article. Clinical Trial.
-
Twisted blood vessels: symptoms, etiology and biomechanical mechanisms.J Vasc Res. 2012;49(3):185-97. doi: 10.1159/000335123. Epub 2012 Mar 14. J Vasc Res. 2012. PMID: 22433458 Free PMC article. Review.
-
Ring Finger Protein 213 in Moyamoya Disease With Pulmonary Arterial Hypertension: A Mini-Review.Front Neurol. 2022 Mar 24;13:843927. doi: 10.3389/fneur.2022.843927. eCollection 2022. Front Neurol. 2022. PMID: 35401401 Free PMC article. Review.
References
-
- Kelleher CM, McLean SE, Mecham RP. Vascular extracellular matrix and aortic development. Curr Top Dev Biol. 2004;62:153–188. - PubMed
-
- Brooke BS, Bayes-Genis A, Li DY. New insights into elastin and vascular disease. Trends Cardiovasc Med. 2003;13:176–181. - PubMed
-
- Pereira L, Andrikopoulos K, Tian J, Lee SY, Keene DR, Ono R, Reinhardt DP, Sakai LY, Biery NJ, Bunton T, Dietz HC, Ramirez F. Targeting of the gene encoding fibrillin-1 recapitulates the vascular aspect of Marfan syndrome. Nat Genet. 1997;17:218–222. - PubMed
-
- Li DY, Brooke B, Davis EC, Mecham RP, Sorensen LK, Boak BB, Eichwald E, Keating MT. Elastin is an essential determinant of arterial morphogenesis. Nature. 1998;393:276–280. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
