

Review
doi: 10.1038/nrc2696.
Emerging roles of E2Fs in cancer: an exit from cell cycle control
Affiliations
- PMID: 19851314
- PMCID: PMC3616489
- DOI: 10.1038/nrc2696
Item in Clipboard
Review
Emerging roles of E2Fs in cancer: an exit from cell cycle control
Nat Rev Cancer.
2009 Nov.
Abstract
Mutations of the retinoblastoma tumour suppressor gene (RB1) or components regulating the RB pathway have been identified in almost every human malignancy. The E2F transcription factors function in cell cycle control and are intimately regulated by RB. Studies of model organisms have revealed conserved functions for E2Fs during development, suggesting that the cancer-related proliferative roles of E2F family members represent a recent evolutionary adaptation. However, given that some human tumours have concurrent RB1 inactivation and E2F amplification and overexpression, we propose that there are alternative tumour-promoting activities for the E2F family, which are independent of cell cycle regulation.
Figures
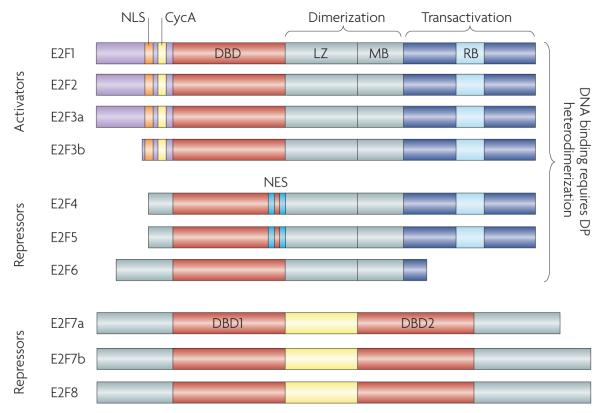
This family is defined by their signature winged-helix DNA binding domain (DBD), and all members are expressed from eight chromosomal loci that encode nine distinct gene products. Based predominantly on results from in vitro studies, the E2F family has been traditionally divided into activator (E2F1–E2F3) and repressor (E2F4–E2F8) subclasses. Most E2F family members (E2F1–E2F6) bind DNA as heterodimers with one of three dimerization partner (DP) proteins, TFDP1, TFDP2 and TFDP3, and heterodimerization is mediated by the leuzine zipper (LZ) and marked box (MB) domains. RB binds within the transactivation domain (RB) of E2F1–E2F3. Alternative promoters at the E2F3 locus drive the expression of two highly related isoforms, E2F3a and E2F3b. Unlike the activating E2F1–E2F3, E2F4 associates with all three pocket protein family members and E2F5 associates predominantly with p130 (REFS 1,2,8). Ectopically expressed E2F activators are localized to the nucleus owing to their amino-terminal nuclear localization signal (NLS) sequence,, which is adjacent to the cyclin A-binding site (CycA). E2F4 and E2F5 have bipartite nuclear export signals (NES) that mediate their export to the cytoplasm,. Repressors E2F6–E2F8 do not possess the canonical carboxy-terminal features of E2F4 and E2F5 and so are presumed to repress E2F-responsive genes independently of RB and related pocket proteins,,,. Indeed, E2F6 can repress E2F target expression when overexpressed in cell culture–. The structurally unique and most recently identified E2Fs, E2F7 and E2F8, comprise a separate and highly evolutionarily conserved repressor arm in the E2F family. Unlike the E2F3 isoforms that are transcribed from distinct promoters, E2F7a and E2F7b isoforms are produced by alternative splicing of the primary transcript.
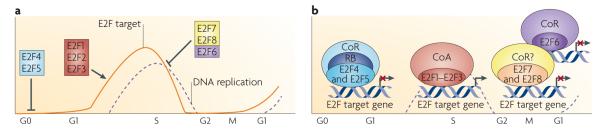
a| In quiescent (G0) cells, the ubiquitously expressed E2F4 and E2F5 associate with pocket proteins and other co-repressors to maintain repression of E2F-responsive genes (line labelled E2F target) that promote entry into the G1 phase of the cell cycle. On mitogenic stimulation, the sequential phosphorylation of RB by activated cyclin-dependent kinases results in the loss of RB function, release of E2F repressors and the accumulation of newly synthesized free E2F1, E2F2 and E2F3 late in G1 phase. Together, these events initiate a transcriptional programme driving cells into S phase (dashed line labelled DNA replication). This G1/S-specific transcriptome is then attenuated on completion of S phase in G2 by the action of the repressors E2F6, E2F7 and E2F8, which are thought to function independently of RB and RB-related proteins,,. E2F7 and E2F8 can also specifically mediate the repression of E2F activators such as E2f1 (REFS 17,41). Heterodimeric or homodimeric E2F7 and E2F8 complexes directly bind the E2f1 promoter and repress its expression throughout S phase to restrain activator E2F function. Without this brake, E2F1 proteins would persist inappropriately and continue the activation of genes encoding components of the transcriptional machinery, resulting in ectopic DNA replication. b| In quiescent or differentiated cells, pocket protein-bound E2F4 and E2F5 have been found to associate with various co-repressors (CoRs), such as histone deacetylases, the DNA methyltransferase DNMT1 and C-terminal binding protein (CtBP), leading to chromatin compaction and transcription inhibition–. Conversely, during cell proliferation, when RB is hyperphosphorylated, E2F activators recruit the basal transcription factor TFIID and other co-activators (CoA), such as histone acetyltransferases, p300 and CBP, GCN5 and TIP60, to specific gene promoters–, leading to an open chromatin configuration and transcription initiation. The more recently identified repressors E2F6–E2F8 mediate repression of E2F-responsive genes independent of pocket protein binding. Although E2F6, through interaction with the polycomb complex, still requires dimerization with a dimerization partner protein to function in transcription repression, E2F7 and E2F8 are unique in that they form homodimers (E2F7–E2F7 and E2F8–E2F8) or heterodimers (E2F7–E2F8) to repress transcription of cell cycle-related genes,,. The co-repressors that associate with E2F7 and E2F8 are currently unknown.
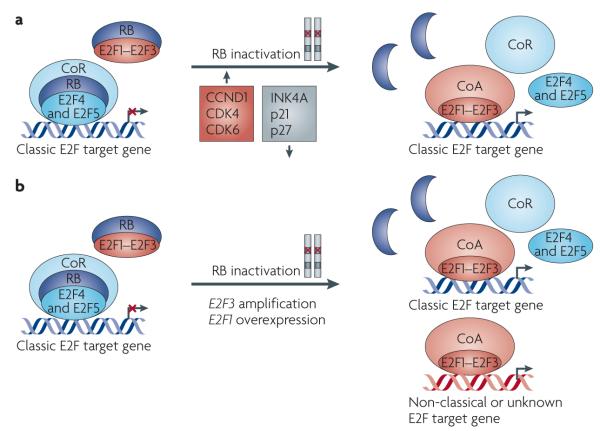
a | In the nuclei of non-proliferating cells, RB remains hypophosphorylated and forms RB–E2F repressor complexes to inhibit the expression of genes that promote S phase entry. Hypophosphorylated RB also directly binds to and inhibits the activity of E2F activators (E2F1–E2F3). RB is inactivated in most types of human cancer and this occurs through the direct mutation of RB1, located on the long arm of chromosome 13 (13q14.3), or through a disruption in the regulatory components of the RB–E2F pathway. For instance, CCND1 is overexpressed or amplified in cancer, an event that leads to deregulated E2F function by activating cyclin-dependent kinase 4 (CDK4) and CDK6 and by stimulating RB hyperphosphorylation. CDK4 and CDK6 are also overexpressed, amplified and mutated in cancers,,, leading to the loss of RB function. Many types of tumours exhibit decreased expression or mutations of CDK inhibitors, such as INK4A, p21 and p27, which normally antagonize RB phosphorylation,,. In human cancer, it is the prevailing view that the inactivation of RB leads to the disassembly of RB–E2F co-repressor complexes, lifting the repression of genes that are necessary for progression through the cell cycle. Furthermore, the dissociation of hyperphosphorylated RB from E2F activators leads to the inappropriate accumulation of free E2F1, E2F2 and E2F3 with unmasked transactivation domains, resulting in additional transactivation of these genes. b | Amplification of the genomic locus 6p22 harbouring E2F3 is detected in more than 50% of cases of retinoblastoma,,, as well as in bladder tumours that have complete loss of RB function–. RB1 is mutated in more than 90% of small-cell lung cancer cases, many of which also exhibit overexpression of E2F1 or E2F3 (REF. 110). The inactivation of RB in these cancers leads to increased proliferation of tumour cells as a result of deregulated E2F function and increased expression of classic E2F target genes. The additional increase in the level of E2F activators might contribute to the activation of genes important for tumour initiation or progression that function independently of cell cycle control. CoA, co-activator; CoR, co-repressor.
References
-
- Trimarchi JM, Lees JA. Sibling rivalry in the E2F family. Nature Rev. Mol. Cell Biol. 2002;3:11–20. - PubMed
-
- Harbour JW, Dean DC. The Rb/E2F pathway: expanding roles and emerging paradigms. Genes Dev. 2000;14:2393–2409. - PubMed
-
- Nevins JR. The Rb/E2F pathway and cancer. Hum. Mol. Genet. 2001;10:699–703. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
