

Review
doi: 10.1038/nrm2780.
Building ubiquitin chains: E2 enzymes at work
Affiliations
- PMID: 19851334
- PMCID: PMC3107738
- DOI: 10.1038/nrm2780
Item in Clipboard
Review
Building ubiquitin chains: E2 enzymes at work
Nat Rev Mol Cell Biol.
2009 Nov.
Abstract
The modification of proteins with ubiquitin chains can change their localization, activity and/or stability. Although ubiquitylation requires the concerted action of ubiquitin-activating enzymes (E1s), ubiquitin-conjugating enzymes (E2s) and ubiquitin ligases (E3s), it is the E2s that have recently emerged as key mediators of chain assembly. These enzymes are able to govern the switch from ubiquitin chain initiation to elongation, regulate the processivity of chain formation and establish the topology of assembled chains, thereby determining the consequences of ubiquitylation for the modified proteins.
Figures
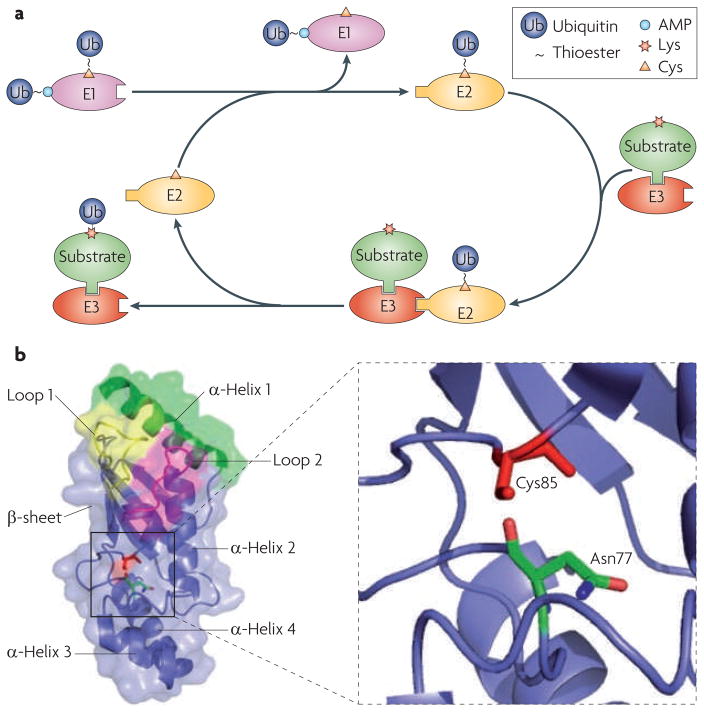
a. Schematic overview of ubiquitylation. A ubiquitin-conjugating enzyme (E2) first interacts with the ubiquitin-activating enzyme (E1) that has been loaded with two ubiquitin molecules (one at its adenylation domain as an adenylate (~AMP) and the other linked to a Cys at its active site as a thioester). The activated ubiquitin is transferred to the Cys in the E2 active site. The E2 has to dissociate from the E1 before it engages with a cognate ubiquitin ligase (E3), which recruits substrates. Once ubiquitin has been transferred to the substrate, the E2 dissociates from the E3, allowing it to be recharged with ubiquitin for the next round of transfers. b. Structure of the core ubiquitin-conjugating (UBC) domain of the E2 UBE2D2. The UBC core is comprised of four α-helices and an antiparallel β-sheet formed by four β–strands. The E1-interacting α-helix 1 is shown in green, and the two E3-interacting loops are shown in yellow (loop 1) and magenta (loop 2). The close-up view shows the relative spatial positions of the side chains of the active-site Cys85 and Asn77.
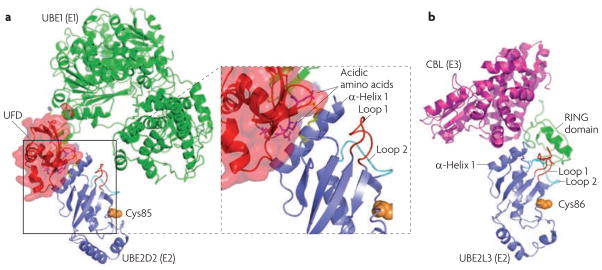
a. Structural model of the ubiquitin-conjugating enzyme (E2)–ubiquitin-activating enzyme (E1) interaction. The example shown is the E2 UBE2D2 in complex with the E1 UBE1 (also| known as UBA1) (Protein Data Bank (PDB) code 3CMM). The structure of UBE2M (the neuronal precursor cell-expressed developmentally downregulated protein 8 (NEDD8) E2) in complex with the ubiquitin fold domain (UFD) of the corresponding E1 is used as a reference (PDB code 1Y8X). The enlarged view shows that the UFD contains a groove formed by acidic amino acids, which are proposed to interact with the side chains of two Lys residues (yellow) in α-helix 1 of the E2. b. The structure of the E2–ubiquitin ligase (E3) interaction. The example shown is the E2 UBE2L3 in complex with the RING E3 casitas B-lineage lymphoma (CBL) (PDB code 1FBV). Note that the E2 has overlapping binding sites on α-helix 1 for the E1 and the E3. The two loops that contact the RING domain and the catalytic Cys86 are also shown.
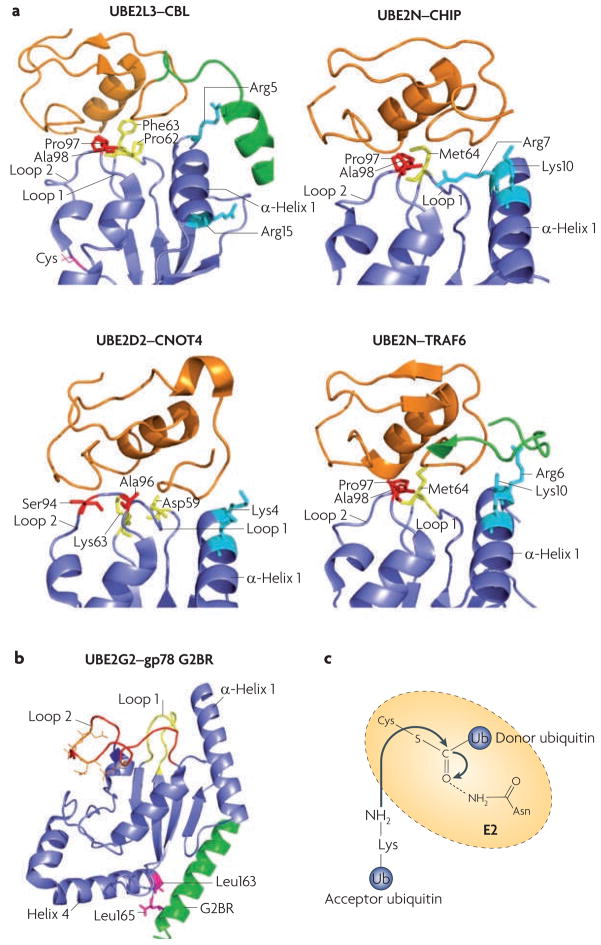
a. Comparison of different ubiquitin-conjugating enzyme (E2)–ubiquitin ligase (E3) interactions: UBE2L3–casitas B-lineage lymphoma (CBL) (Protein Data Bank (PDB) code 1FBV), UBE2N–carboxy terminus of HSC70 interacting protein (CHIP; also known as STUB1) (PDB code 2C2V), UBE2D2–CNOT4 (CCR4–NOT transcription complex, subunit 4) (PDB code 1UR6) and UBE2N–TNF receptor associated factor 6 (TRAF6) (PDB code 3HCT). All E2s use loop 1, loop 2 and α-helix 1 to interact with E3s. The side chains of the residues involved in E3 recognition are colour-coded (residues in loop 1 are yellow, in loop 2 are red and in α-helix 1 are cyan). Note that for CBL and TRAF6, E3 elements outside the RING domain (shown in green) participate in E2 binding. The CNOT4–UE2D2 structure is an NMR model, whereas the others were solved by crystallography. The RING finger (or the U-Box for CHIP) is in orange, and the cognate E2s are in blue. b. An example structure of a RING-independent E2–E3 interaction: that between UBE2G2 (blue) and the G2BR peptide from gp78 (also known as AMFR; green) (PBD code 3FSH). The E3 peptide interacts with two Leu residues of the E2, at a position remote from the predicted RING binding site (loop 1, loop 2 and α-helix 1). Acidic residues in the UBE2G2 acidic loop are in orange. c. A catalysis model for isopeptide bond formation during ubiquitin chain synthesis. The side chain of a conserved Asn residue in the E2 is proposed to interact with the active-site Cys (carrying the donor ubiquitin), which stabilizes the oxyanion transition state of the nucleophilic attack by the Lys residue of the acceptor ubiquitin.
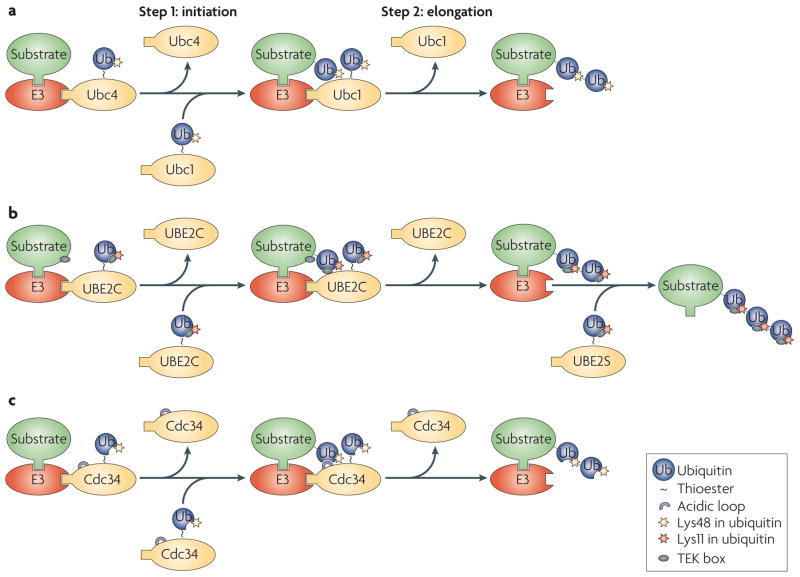
a. The yeast anaphase-promoting complex or cyclosome (APC/C) uses the ubiquitin-conjugating enzyme (E2) Ubc4 to initiate ubiquitylation, and a different E2, Ubc1, to elongate Lys48-linked ubiquitin chains. b. UBE2C recognizes a TEK box in the substrate to initiate ubiquitylation. Once a ubiquitin moiety has been added to the substrate, a similar TEK box present in ubiquitin takes over to promote elongation of the Lys11-linked ubiquitin chain. The efficient extension of Lys11-linked ubiquitin chains requires a second E2, UBE2S. c. The yeast E2 cell division cycle 34 (Cdc34) can initiate and elongate ubiquitin chains. Efficient chain elongation requires the acidic loop of Cdc34, which might orient the attacking Lys group relative to the E2 active site (charged with the ubiquitin thioester) to facilitate the formation of ubiquitin–ubiquitin linkage. E3, ubiquitin ligase.
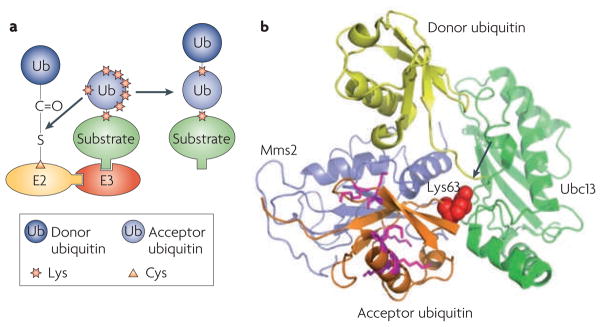
a. A model for ubiquitin chain linkage selection by ubiquitin-conjugating enzymes (E2s). The E2 orients the acceptor ubiquitin in a way that exposes only the favoured Lys residue to its active site (charged with the donor ubiquitin), leading to the formation of ubiquitin chains of a specific linkage. b. Structural illustration of ubiquitin chain linkage selection by the E2 heterodimer Ubc13–Mms2 (Protein Data Bank code 2GMI). In the structure of the Ubc13~Ub-Mms2 complex (where ~ represents a thioester bond), an acceptor ubiquitin from a neighbouring complex in the crystal makes contact with Mms2 in such a way that only its Lys63 is aligned with the thioester (arrow) that links Ubc13 to the donor ubiquitin.
References
-
- Kerscher O, Felberbaum R, Hochstrasser M. Modification of proteins by ubiquitin and ubiquitin-like proteins. Annu Rev Cell Dev Biol. 2006;22:159–180. - PubMed
-
- Mukhopadhyay D, Riezman H. Proteasome-independent functions of ubiquitin in endocytosis and signaling. Science. 2007;315:201–205. - PubMed
-
- Pickart CM, Fushman D. Polyubiquitin chains: polymeric protein signals. Curr Opin Chem Biol. 2004;8:610–616. - PubMed
-
- Tokunaga F, et al. Involvement of linear polyubiquitylation of NEMO in NF-κB activation. Nature Cell Biol. 2009;11:123–132. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
