

Loss of yeast peroxiredoxin Tsa1p induces genome instability through activation of the DNA damage checkpoint and elevation of dNTP levels
- PMID: 19851444
- PMCID: PMC2758587
- DOI: 10.1371/journal.pgen.1000697
Loss of yeast peroxiredoxin Tsa1p induces genome instability through activation of the DNA damage checkpoint and elevation of dNTP levels
Abstract
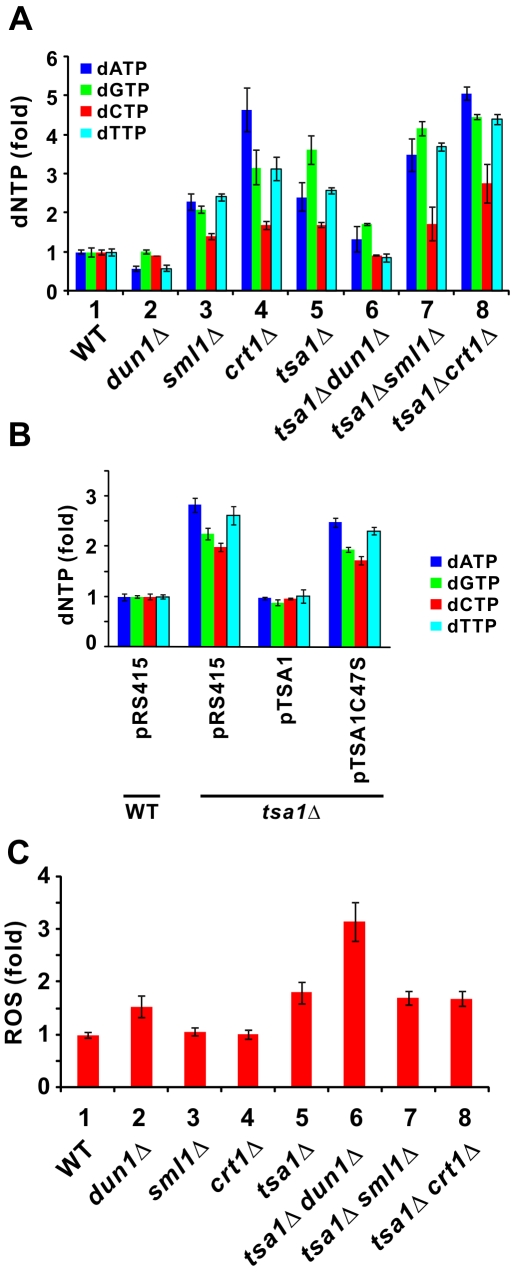
Peroxiredoxins are a family of antioxidant enzymes critically involved in cellular defense and signaling. Particularly, yeast peroxiredoxin Tsa1p is thought to play a role in the maintenance of genome integrity, but the underlying mechanism is not understood. In this study, we took a genetic approach to investigate the cause of genome instability in tsa1Delta cells. Strong genetic interactions of TSA1 with DNA damage checkpoint components DUN1, SML1, and CRT1 were found when mutant cells were analyzed for either sensitivity to DNA damage or rate of spontaneous base substitutions. An elevation in intracellular dNTP production was observed in tsa1Delta cells. This was associated with constitutive activation of the DNA damage checkpoint as indicated by phosphorylation of Rad9/Rad53p, reduced steady-state amount of Sml1p, and induction of RNR and HUG1 genes. In addition, defects in the DNA damage checkpoint did not modulate intracellular level of reactive oxygen species, but suppressed the mutator phenotype of tsa1Delta cells. On the contrary, overexpression of RNR1 exacerbated this phenotype by increasing dNTP levels. Taken together, our findings uncover a new role of TSA1 in preventing the overproduction of dNTPs, which is a root cause of genome instability.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
-
- Rhee SG, Chae HZ, Kim K. Peroxiredoxins: a historical overview and speculative preview of novel mechanisms and emerging concepts in cell signaling. Free Radic Biol Med. 2005;38:1543–1552. - PubMed
-
- Fourquet S, Huang ME, D'Autreaux B, Toledano MB. The dual functions of thiol-based peroxidases in H2O2 scavenging and signaling. Antioxid Redox Signal. 2008;10:1565–1576. - PubMed
-
- Bryk R, Griffin P, Nathan C. Peroxynitrite reductase activity of bacterial peroxiredoxins. Nature. 2000;407:211–215. - PubMed
-
- Wong CM, Zhou Y, Ng RWM, Kung HF, Jin DY. Cooperation of yeast peroxiredoxins Tsa1p and Tsa2p in the cellular defense against oxidative and nitrosative stress. J Biol Chem. 2002;277:5385–5394. - PubMed
-
- Jin DY, Chae HZ, Rhee SG, Jeang KT. Regulatory role for a novel human thioredoxin peroxidase in NF-κB activation. J Biol Chem. 1997;272:30952–30961. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
