

Minireview: the melanocortin 2 receptor accessory proteins
- PMID: 19855089
- PMCID: PMC5419097
- DOI: 10.1210/me.2009-0283
Minireview: the melanocortin 2 receptor accessory proteins
Abstract
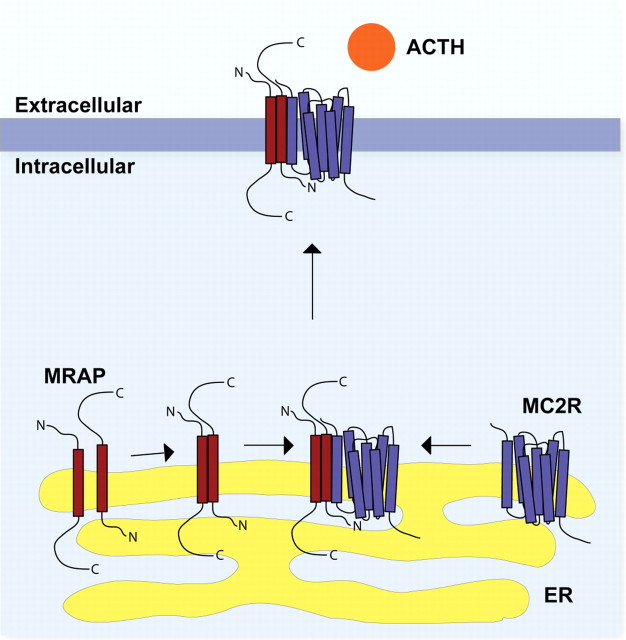
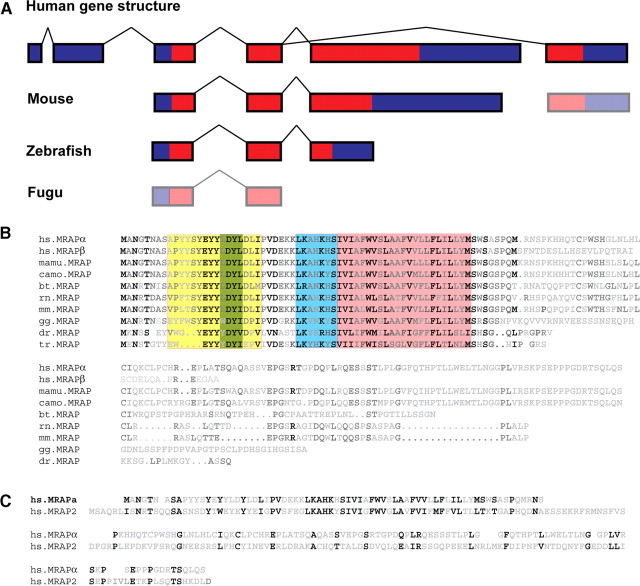
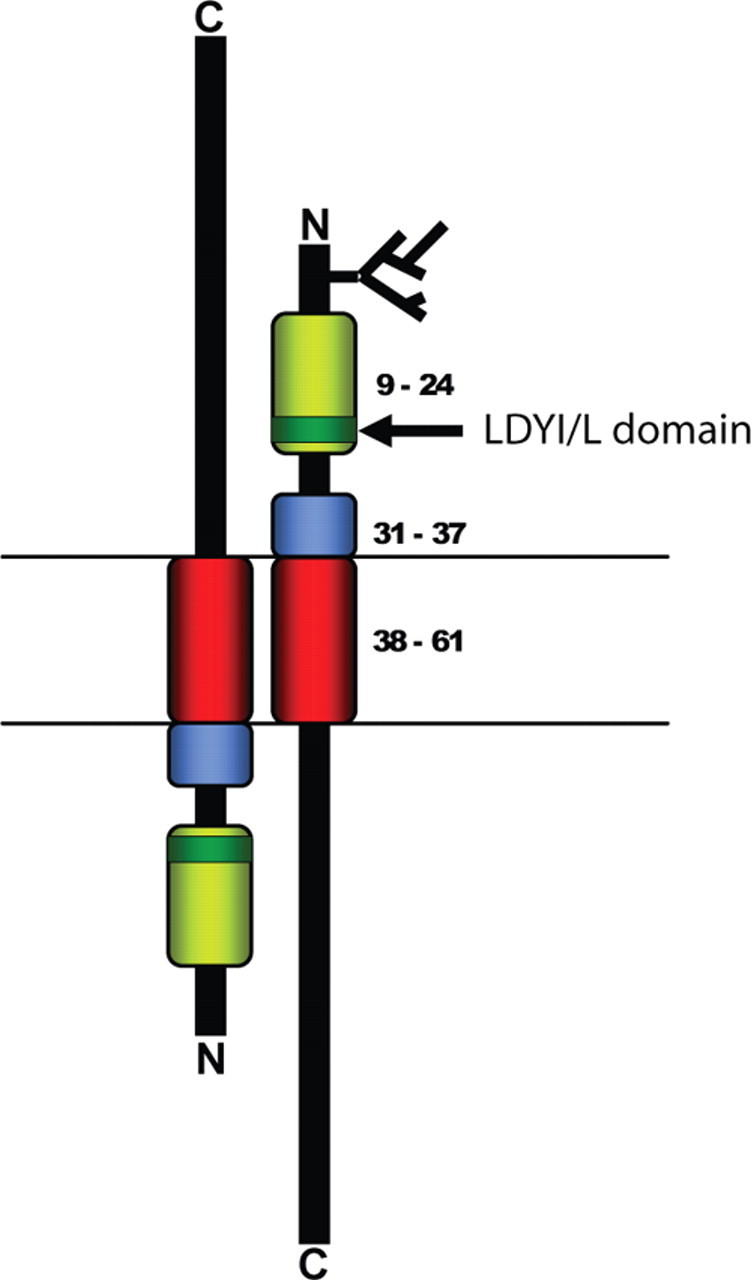
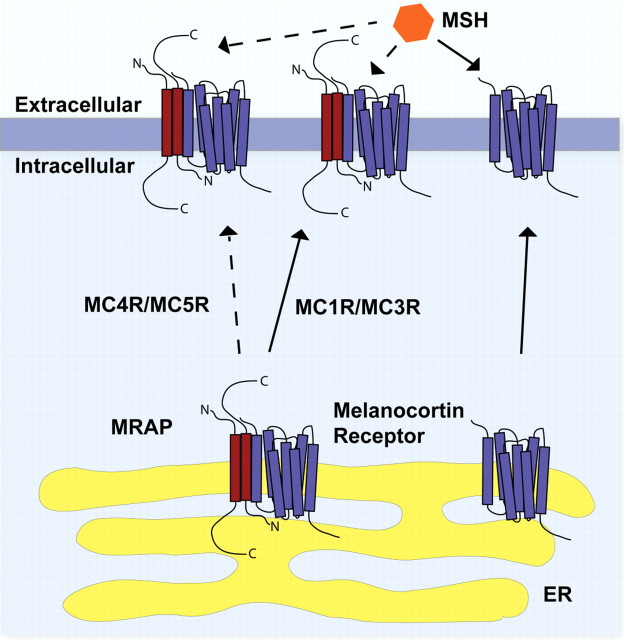
The melanocortin 2 receptor (MC2R) accessory protein, MRAP, is one of a growing number of G protein-coupled receptor accessory proteins that have been identified in recent years that add control and complexity to G protein-coupled receptor functional expression and signal transduction. MRAP interacts directly with MC2R and is essential for its trafficking from the endoplasmic reticulum to the cell surface, where it acts as the receptor for the pituitary hormone ACTH. In addition, MRAP2, a newly described homolog of MRAP, is also able to support the cell surface expression of MC2R. Although it is clear that MRAP is required for MC2R function, the mechanism of MRAP action is only beginning to be understood. Recent work has started to reveal some of these mechanisms and the MRAP domains involved in MC2R functional expression, and new data have shown a potential role for both MRAP and MRAP2 in the regulation of the other melanocortin receptors.
Figures

References
-
- Metherell LA, Chapple JP, Cooray S, David A, Becker C, Rüschendorf F, Naville D, Begeot M, Khoo B, Nürnberg P, Huebner A, Cheetham ME, Clark AJL2005. Mutations in MRAP, encoding a new interacting partner of the ACTH receptor, cause familial glucocorticoid deficiency type 2. Nat Genet 37:166–170 - PubMed
-
- Cone RD, Mountjoy KG, Robbins LS Nadeau JH, Johnson KR, Roselli-Rehfuss L, Mortrud MT1993. Cloning and functional characterization of a family of receptors for the melanotropic peptides. Ann NY Acad Sci 680:342–363 - PubMed
-
- Mountjoy KG, Robbins LS, Mortrud MT, Cone RD1992. The cloning of a family of genes that encode the melanocortin receptors. Science 257:1248–1251 - PubMed
-
- Cone RD2006. Studies on the physiological functions of the melanocortin system. Endocr Rev 27:736–749 - PubMed
-
- Noon LA, Franklin JM, King PJ, Goulding NJ, Hunyady L, Clark AJ2002. Failed export of the adrenocorticotrophin receptor from the endoplasmic reticulum in non-adrenal cells: evidence in support of a requirement for a specific adrenal accessory factor. J Endocrinol 174:17–25 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
