

Loading-related regulation of gene expression in bone in the contexts of estrogen deficiency, lack of estrogen receptor alpha and disuse
- PMID: 19857613
- PMCID: PMC2887492
- DOI: 10.1016/j.bone.2009.10.021
Loading-related regulation of gene expression in bone in the contexts of estrogen deficiency, lack of estrogen receptor alpha and disuse
Abstract
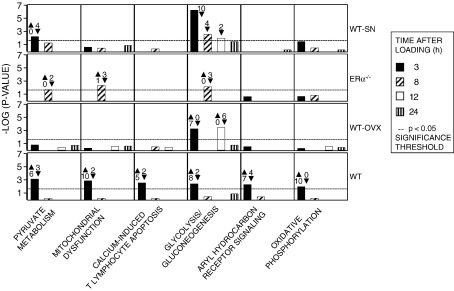
Loading-related changes in gene expression in resident cells in the tibia of female mice in the contexts of normality (WT), estrogen deficiency (WT-OVX), absence of estrogen receptor alpha (ERalpha(-/-)) and disuse due to sciatic neurectomy (WT-SN) were established by microarray. Total RNA was extracted from loaded and contra-lateral non-loaded tibiae at selected time points after a single, short period of dynamic loading sufficient to engender an osteogenic response. There were marked changes in the expression of many genes according to context as well as in response to loading within those contexts. In WT mice at 3, 8, 12 and 24 h after loading the expression of 642, 341, 171 and 24 genes, respectively, were differentially regulated compared with contra-lateral bones which were not loaded. Only a few of the genes differentially regulated by loading in the tibiae of WT mice have recognized roles in bone metabolism or have been linked previously to osteogenesis (Opn, Sost, Esr1, Tgfb1, Lrp1, Ostn, Timp, Mmp, Ctgf, Postn and Irs1, BMP and DLX5). The canonical pathways showing the greatest loading-related regulation were those involving pyruvate metabolism, mitochondrial dysfunction, calcium-induced apoptosis, glycolysis/gluconeogenesis, aryl hydrocarbon receptor and oxidative phosphorylation. In the tibiae from WT-OVX, ERalpha(-/-) and WT-SN mice, 440, 439 and 987 genes respectively were differentially regulated by context alone compared to WT. The early response to loading in tibiae of WT-OVX mice involved differential regulation compared to their contra-lateral non-loaded pair of fewer genes than in WT, more down-regulation than up-regulation and a later response. This was shared by WT-SN. In tibiae of ERalpha(-/-) mice, the number of genes differentially regulated by loading was markedly reduced at all time points. These data indicate that in resident bone cells, both basal and loading-related gene expression is substantially modified by context. Many of the genes differentially regulated by the earliest loading-related response were primarily involved in energy metabolism and were not specific to bone.
Figures







References
-
- Frost H.M. The mechanostat: a proposed pathogenic mechanism of osteoporoses and the bone mass effects of mechanical and nonmechanical agents. Bone Miner. 1987;2:73–85. - PubMed
-
- Wolf J. Das Gesetz der Transformation der Knochen. Berlin: A Hirschwald [trans. P Maquet & R Furlong as The Law of Bone Remodelling. Berlin: Springer, 1986] 1892.
-
- Welten D.C., Kemper H.C., Post G.B., Van Mechelen W., Twisk J., Lips P. Weight-bearing activity during youth is a more important factor for peak bone mass than calcium intake. J. Bone Miner. Res. 1994;9:1089–1096. - PubMed
-
- Pead M.J., Skerry T.M., Lanyon L.E. Direct transformation from quiescence to bone formation in the adult periosteum following a single brief period of bone loading. J. Bone Miner. Res. 1988;3:647–656. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
