

Mapping simocyclinone D8 interaction with DNA gyrase: evidence for a new binding site on GyrB
- PMID: 19858260
- PMCID: PMC2798491
- DOI: 10.1128/AAC.00972-09
Mapping simocyclinone D8 interaction with DNA gyrase: evidence for a new binding site on GyrB
Abstract
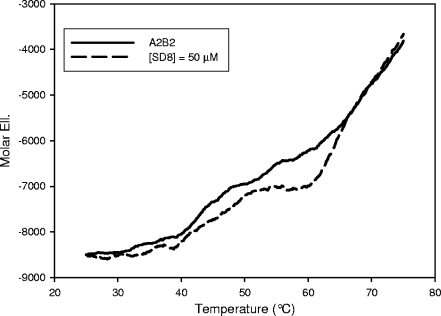
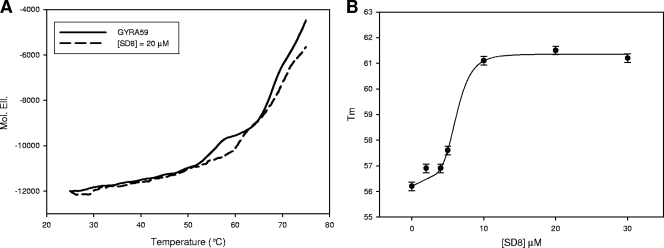
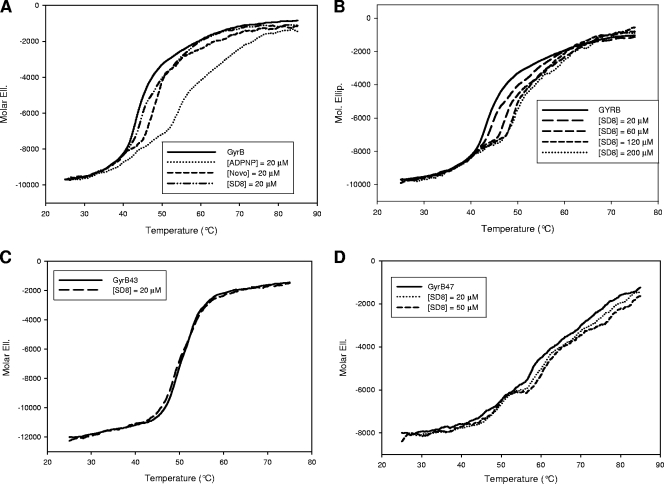
Simocyclinone D8, a coumarin derivative isolated from Streptomyces antibioticus Tü 6040, represents an interesting new antiproliferative agent. It was originally suggested that this drug recognizes the GyrA subunit and interferes with the gyrase catalytic cycle by preventing its binding to DNA. To further characterize the mode of action of this antibiotic, we investigated its binding to the reconstituted DNA gyrase (A(2)B(2)) as well as to its GyrA and GyrB subunits and the individual domains of these proteins, by performing protein melting and proteolytic digestion studies as well as inhibition assays. Two binding sites were identified, one (anticipated) in the N-terminal domain of GyrA (GyrA59) and the other (unexpected) at the C-terminal domain of GyrB (GyrB47). Stabilization of the A subunit appears to be considerably more effective than stabilization of the B subunit. Our data suggest that these two distinct sites could cooperate in the reconstituted enzyme.
Figures
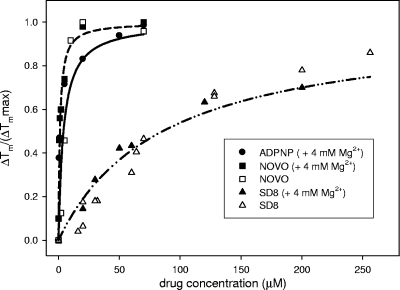
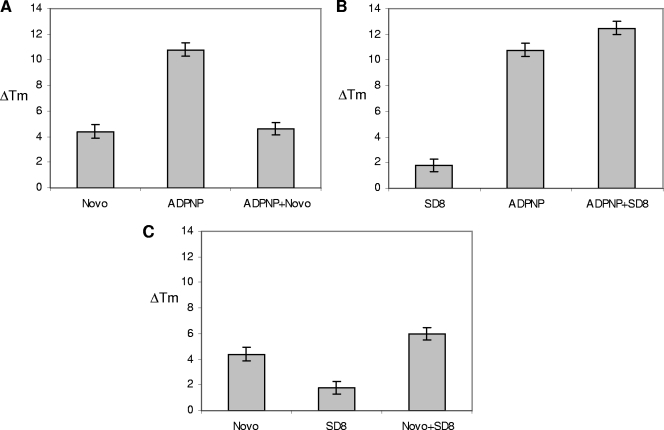
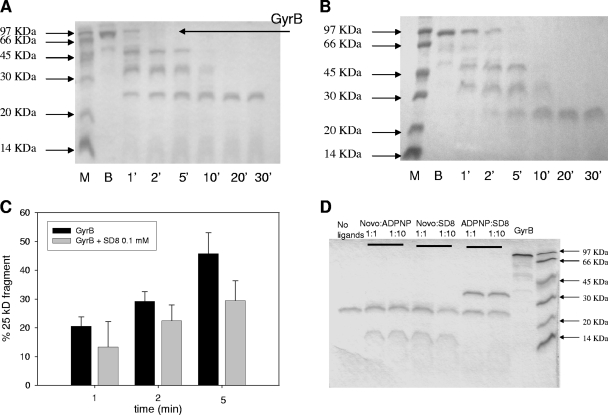
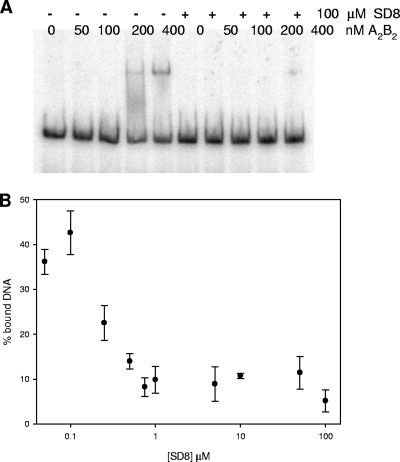
Similar articles
-
Simocyclinone D8, an inhibitor of DNA gyrase with a novel mode of action.Antimicrob Agents Chemother. 2005 Mar;49(3):1093-100. doi: 10.1128/AAC.49.3.1093-1100.2005. Antimicrob Agents Chemother. 2005. PMID: 15728908 Free PMC article.
-
A new crystal structure of the bifunctional antibiotic simocyclinone D8 bound to DNA gyrase gives fresh insight into the mechanism of inhibition.J Mol Biol. 2014 May 15;426(10):2023-33. doi: 10.1016/j.jmb.2014.02.017. Epub 2014 Mar 1. J Mol Biol. 2014. PMID: 24594357 Free PMC article.
-
A crystal structure of the bifunctional antibiotic simocyclinone D8, bound to DNA gyrase.Science. 2009 Dec 4;326(5958):1415-8. doi: 10.1126/science.1179123. Science. 2009. PMID: 19965760
-
Discovery and development of ATPase inhibitors of DNA gyrase as antibacterial agents.Curr Med Chem. 2007;14(19):2033-47. doi: 10.2174/092986707781368414. Curr Med Chem. 2007. PMID: 17691945 Review.
-
The interaction between coumarin drugs and DNA gyrase.Mol Microbiol. 1993 Aug;9(4):681-6. doi: 10.1111/j.1365-2958.1993.tb01728.x. Mol Microbiol. 1993. PMID: 8231802 Review.
Cited by
-
Identification of novel bacterial DNA gyrase inhibitors: An in silico study.Res Pharm Sci. 2016 May-Jun;11(3):250-8. Res Pharm Sci. 2016. PMID: 27499795 Free PMC article.
-
SimReg1 is a master switch for biosynthesis and export of simocyclinone D8 and its precursors.AMB Express. 2012 Jan 3;2(1):1. doi: 10.1186/2191-0855-2-1. AMB Express. 2012. PMID: 22214346 Free PMC article.
-
Topoisomerases as anticancer targets.Biochem J. 2018 Jan 23;475(2):373-398. doi: 10.1042/BCJ20160583. Biochem J. 2018. PMID: 29363591 Free PMC article. Review.
-
Protective effect of Qnr on agents other than quinolones that target DNA gyrase.Antimicrob Agents Chemother. 2015 Nov;59(11):6689-95. doi: 10.1128/AAC.01292-15. Epub 2015 Aug 3. Antimicrob Agents Chemother. 2015. PMID: 26239981 Free PMC article.
-
In front of and behind the replication fork: bacterial type IIA topoisomerases.Cell Mol Life Sci. 2010 Jun;67(12):2001-24. doi: 10.1007/s00018-010-0299-5. Epub 2010 Feb 18. Cell Mol Life Sci. 2010. PMID: 20165898 Free PMC article. Review.
References
-
- Ali, J. A., A. P. Jackson, A. J. Howells, and A. Maxwell. 1993. The 43-kilodalton N-terminal fragment of the DNA gyrase B protein hydrolyzes ATP and binds coumarin drugs. Biochemistry 32:2717-2724. - PubMed
-
- Ali, J. A., G. Orphanides, and A. Maxwell. 1995. Nucleotide binding to the 43-kilodalton N-terminal fragment of the DNA gyrase B protein. Biochemistry 34:9801-9808. - PubMed
-
- Appelbaum, P. C., and P. A. Hunter. 2000. The fluoroquinolone antibacterials: past, present and future perspectives. Int. J. Antimicrob. Agents 16:5-15. - PubMed
-
- Blandamer, M. J., B. Briggs, P. M. Cullis, A. P. Jackson, A. Maxwell, and R. J. Reece. 1994. Domain structure of Escherichia coli DNA gyrase as revealed by differential scanning calorimetry. Biochemistry 33:7510-7516. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
