

Affinity for phosphatidylinositol 4,5-bisphosphate determines muscarinic agonist sensitivity of Kv7 K+ channels
- PMID: 19858360
- PMCID: PMC2768799
- DOI: 10.1085/jgp.200910313
Affinity for phosphatidylinositol 4,5-bisphosphate determines muscarinic agonist sensitivity of Kv7 K+ channels
Abstract
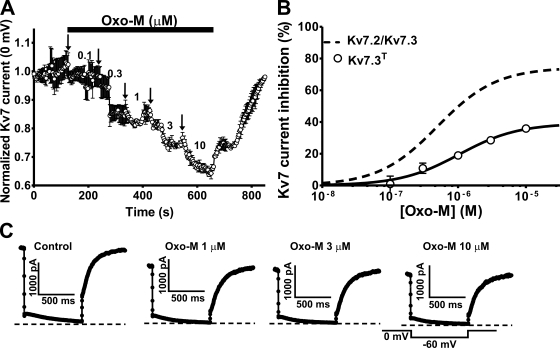
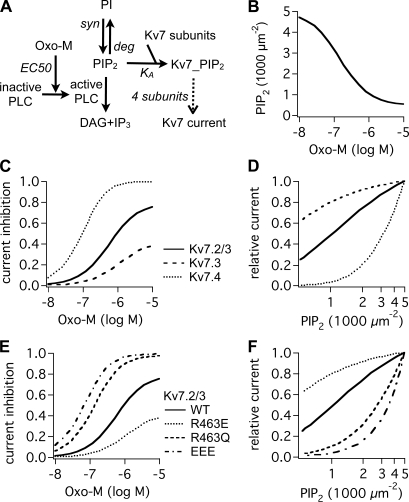
Kv7 K(+)-channel subunits differ in their apparent affinity for PIP(2) and are differentially expressed in nerve, muscle, and epithelia in accord with their physiological roles in those tissues. To investigate how PIP(2) affinity affects the response to physiological stimuli such as receptor stimulation, we exposed homomeric and heteromeric Kv7.2, 7.3, and 7.4 channels to a range of concentrations of the muscarinic receptor agonist oxotremorine-M (oxo-M) in a heterologous expression system. Activation of M(1) receptors by oxo-M leads to PIP(2) depletion through G(q) and phospholipase C (PLC). Chinese hamster ovary cells were transiently transfected with Kv7 subunits and M(1) receptors and studied under perforated-patch voltage clamp. For Kv7.2/7.3 heteromers, the EC(50) for current suppression was 0.44 +/- 0.08 microM, and the maximal inhibition (Inhib(max)) was 74 +/- 3% (n = 5-7). When tonic PIP(2) abundance was increased by overexpression of PIP 5-kinase, the EC(50) was shifted threefold to the right (1.2 +/- 0.1 microM), but without a significant change in Inhib(max) (73 +/- 4%, n = 5). To investigate the muscarinic sensitivity of Kv7.3 homomers, we used the A315T pore mutant (Kv7.3(T)) that increases whole-cell currents by 30-fold without any change in apparent PIP(2) affinity. Kv7.3(T) currents had a slightly right-shifted EC(50) as compared with Kv7.2/7.3 heteromers (1.0 +/- 0.8 microM) and a strongly reduced Inhib(max) (39 +/- 3%). In contrast, the dose-response curve of homomeric Kv7.4 channels was shifted considerably to the left (66 +/- 8 nM), and Inhib(max) was slightly increased (81 +/- 6%, n = 3-4). We then studied several Kv7.2 mutants with altered apparent affinities for PIP(2) by coexpressing them with Kv7.3(T) subunits to boost current amplitudes. For the lower affinity (Kv7.2 (R463Q)/Kv7.3(T)) or higher affinity (Kv7.2 (R463E)/Kv7.3(T)) channels, the EC(50) and Inhib(max) were similar to Kv7.4 or Kv7.3(T) homomers (0.12 +/- 0.08 microM and 79 +/- 6% [n = 3-4] and 0.58 +/- 0.07 microM and 27 +/- 3% [n = 3-4], respectively). The very low-affinity Kv7.2 (R452E, R459E, and R461E) triple mutant was also coexpressed with Kv7.3(T). The resulting heteromer displayed a very low EC(50) for inhibition (32 +/- 8 nM) and a slightly increased Inhib(max) (83 +/- 3%, n = 3-4). We then constructed a cellular model that incorporates PLC activation by oxo-M, PIP(2) hydrolysis, PIP(2) binding to Kv7-channel subunits, and K(+) current through Kv7 tetramers. We were able to fully reproduce our data and extract a consistent set of PIP(2) affinities.
Figures



Similar articles
-
Distinct subunit contributions to the activation of M-type potassium channels by PI(4,5)P2.J Gen Physiol. 2012 Jul;140(1):41-53. doi: 10.1085/jgp.201210796. Epub 2012 Jun 11. J Gen Physiol. 2012. PMID: 22689829 Free PMC article.
-
Regulation of Kv7 (KCNQ) K+ channel open probability by phosphatidylinositol 4,5-bisphosphate.J Neurosci. 2005 Oct 26;25(43):9825-35. doi: 10.1523/JNEUROSCI.2597-05.2005. J Neurosci. 2005. PMID: 16251430 Free PMC article.
-
Angiotensin II regulates neuronal excitability via phosphatidylinositol 4,5-bisphosphate-dependent modulation of Kv7 (M-type) K+ channels.J Physiol. 2006 Aug 15;575(Pt 1):49-67. doi: 10.1113/jphysiol.2006.114074. Epub 2006 Jun 15. J Physiol. 2006. PMID: 16777936 Free PMC article.
-
Neural KCNQ (Kv7) channels.Br J Pharmacol. 2009 Apr;156(8):1185-95. doi: 10.1111/j.1476-5381.2009.00111.x. Epub 2009 Mar 9. Br J Pharmacol. 2009. PMID: 19298256 Free PMC article. Review.
-
Regulation of neural KCNQ channels: signalling pathways, structural motifs and functional implications.J Physiol. 2008 Apr 1;586(7):1811-21. doi: 10.1113/jphysiol.2007.148304. Epub 2008 Jan 31. J Physiol. 2008. PMID: 18238808 Free PMC article. Review.
Cited by
-
The calcium-activated slow AHP: cutting through the Gordian knot.Front Cell Neurosci. 2012 Oct 25;6:47. doi: 10.3389/fncel.2012.00047. eCollection 2012. Front Cell Neurosci. 2012. PMID: 23112761 Free PMC article.
-
Potent KCNQ2/3-specific channel activator suppresses in vivo epileptic activity and prevents the development of tinnitus.J Neurosci. 2015 Jun 10;35(23):8829-42. doi: 10.1523/JNEUROSCI.5176-14.2015. J Neurosci. 2015. PMID: 26063916 Free PMC article.
-
Distinct subunit contributions to the activation of M-type potassium channels by PI(4,5)P2.J Gen Physiol. 2012 Jul;140(1):41-53. doi: 10.1085/jgp.201210796. Epub 2012 Jun 11. J Gen Physiol. 2012. PMID: 22689829 Free PMC article.
-
Triggering actin comets versus membrane ruffles: distinctive effects of phosphoinositides on actin reorganization.Sci Signal. 2011 Dec 13;4(203):ra87. doi: 10.1126/scisignal.2002033. Sci Signal. 2011. PMID: 22169478 Free PMC article.
-
Gq-Coupled Muscarinic Receptor Enhancement of KCNQ2/3 Channels and Activation of TRPC Channels in Multimodal Control of Excitability in Dentate Gyrus Granule Cells.J Neurosci. 2019 Feb 27;39(9):1566-1587. doi: 10.1523/JNEUROSCI.1781-18.2018. Epub 2018 Dec 28. J Neurosci. 2019. PMID: 30593498 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
