

Extending the transposable payload limit of Sleeping Beauty (SB) using the Herpes Simplex Virus (HSV)/SB amplicon-vector platform
- PMID: 19865178
- PMCID: PMC2837128
- DOI: 10.1038/gt.2009.144
Extending the transposable payload limit of Sleeping Beauty (SB) using the Herpes Simplex Virus (HSV)/SB amplicon-vector platform
Abstract
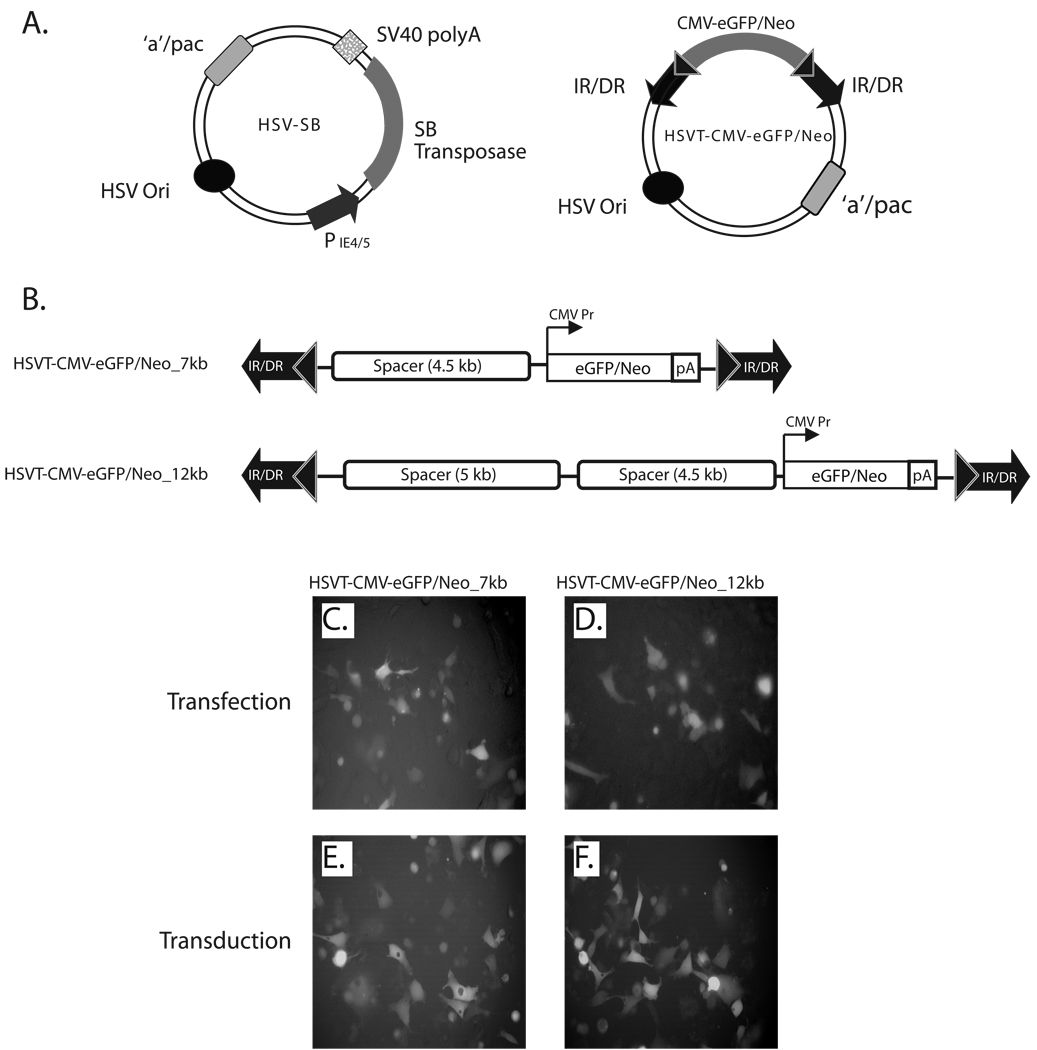
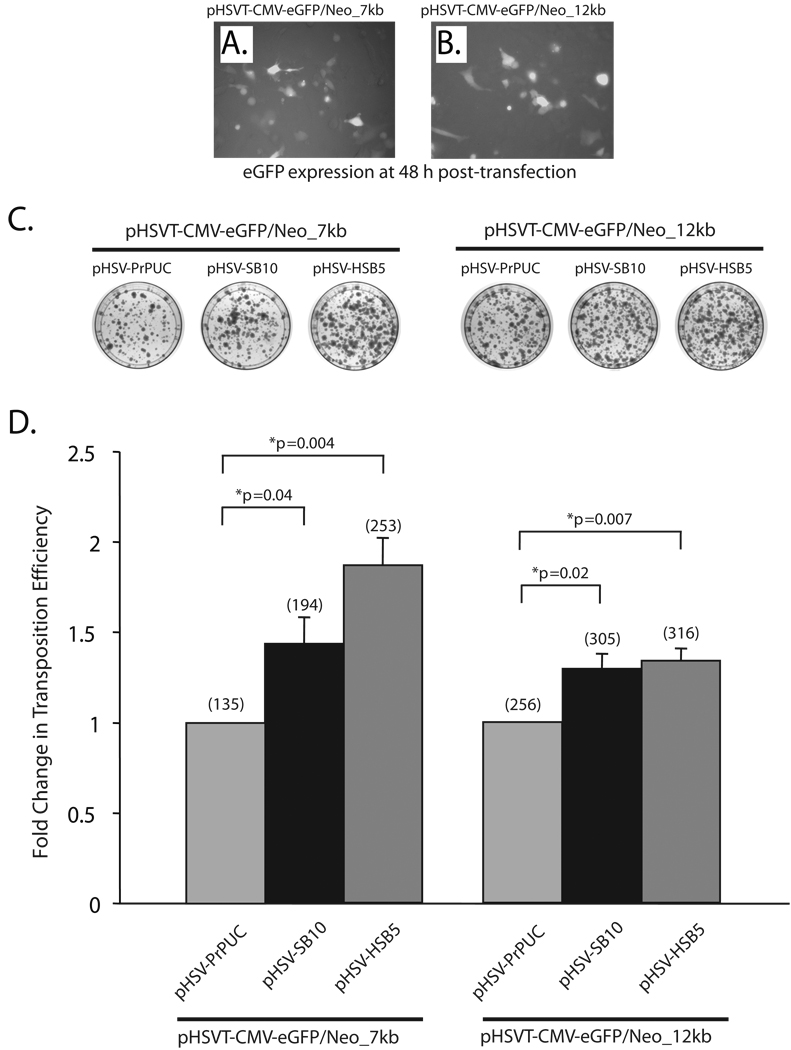
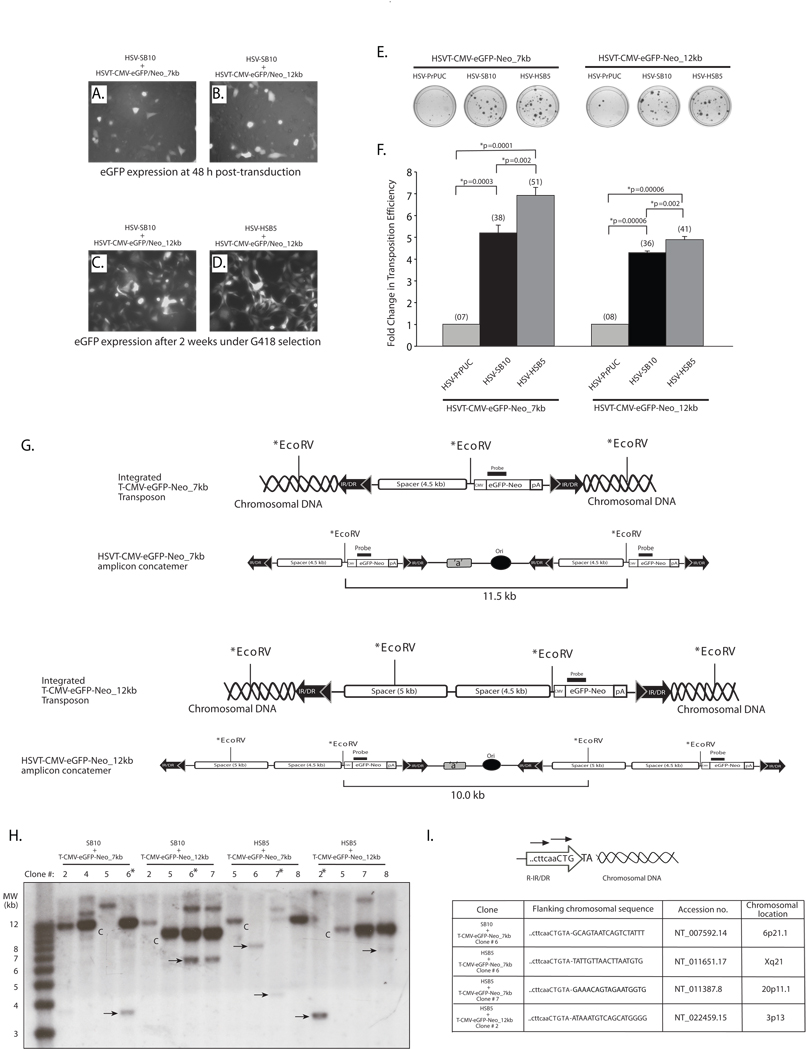
The ability of a viral vector to safely deliver and stably integrate large transgene units (transgenons), which not only include one or several therapeutic genes, but also requisite native transcriptional regulatory elements, would be of significant benefit for diseases presently refractory to available technologies. The herpes simplex virus type-1 (HSV-1) amplicon vector has the largest known payload capacity of approximately 130 kb, but its episomal maintenance within the transduced cell nucleus and induction of host cell silencing mechanisms limits the duration of the delivered therapeutic gene(s). Our laboratory developed an integration-competent version of the HSV-1 amplicon by adaptation of the Sleeping Beauty (SB) transposon system, which significantly extends transgene expression in vivo. The maximum size limit of the amplicon-vectored transposable element remains unknown, but previously published plasmid-centric studies have established that DNA segments longer than 6-kb are inefficiently transposed. Here, we compared the transposition efficiency of SB transposase in the context of both the HSV amplicon vector as well as the HSV amplicon plasmid harboring 7 and 12-kb transposable reporter transgene units. Our results indicate that the transposition efficiency of the 12-kb transposable unit via SB transposase was significantly reduced as compared with the 7-kb transposable unit when the plasmid version of the HSV amplicon was used. However, the packaged HSV amplicon vector form provided a more amenable platform from which the 12-kb transposable unit was mobilized at efficiency similar to that of the 7-kb transposable unit via the SB transposase. Overall, our results indicate that SB is competent in stably integrating transgenon units of at least 12 kb in size within the human genome upon delivery of the platform via HSV amplicons.
Figures
Similar articles
-
Virion-associated cofactor high-mobility group DNA-binding protein-1 facilitates transposition from the herpes simplex virus/Sleeping Beauty amplicon vector platform.Hum Gene Ther. 2010 Nov;21(11):1615-22. doi: 10.1089/hum.2010.022. Epub 2010 Oct 7. Hum Gene Ther. 2010. PMID: 20568967 Free PMC article.
-
Herpes simplex virus/Sleeping Beauty vector-based embryonic gene transfer using the HSB5 mutant: loss of apparent transposition hyperactivity in vivo.Hum Gene Ther. 2010 Nov;21(11):1603-13. doi: 10.1089/hum.2010.062. Epub 2010 Oct 22. Hum Gene Ther. 2010. PMID: 20507234 Free PMC article.
-
Hybrid adeno-associated viral vectors utilizing transposase-mediated somatic integration for stable transgene expression in human cells.PLoS One. 2013 Oct 8;8(10):e76771. doi: 10.1371/journal.pone.0076771. eCollection 2013. PLoS One. 2013. PMID: 24116154 Free PMC article.
-
Herpes simplex virus type 1 amplicons and their hybrid virus partners, EBV, AAV, and retrovirus.Curr Gene Ther. 2004 Dec;4(4):385-408. doi: 10.2174/1566523043346129. Curr Gene Ther. 2004. PMID: 15578989 Review.
-
Development of adenovirus hybrid vectors for Sleeping Beauty transposition in large mammals.Curr Gene Ther. 2011 Oct;11(5):363-74. doi: 10.2174/156652311797415890. Curr Gene Ther. 2011. PMID: 21888620 Review.
Cited by
-
Translating Sleeping Beauty transposition into cellular therapies: victories and challenges.Bioessays. 2010 Sep;32(9):756-67. doi: 10.1002/bies.201000027. Bioessays. 2010. PMID: 20652893 Free PMC article. Review.
-
Virion-associated cofactor high-mobility group DNA-binding protein-1 facilitates transposition from the herpes simplex virus/Sleeping Beauty amplicon vector platform.Hum Gene Ther. 2010 Nov;21(11):1615-22. doi: 10.1089/hum.2010.022. Epub 2010 Oct 7. Hum Gene Ther. 2010. PMID: 20568967 Free PMC article.
-
Preclinical and clinical advances in transposon-based gene therapy.Biosci Rep. 2017 Dec 5;37(6):BSR20160614. doi: 10.1042/BSR20160614. Print 2017 Dec 22. Biosci Rep. 2017. PMID: 29089466 Free PMC article. Review.
-
Targeting the central nervous system with herpes simplex virus / Sleeping Beauty hybrid amplicon vectors.Curr Gene Ther. 2011 Oct;11(5):332-40. doi: 10.2174/156652311797415845. Curr Gene Ther. 2011. PMID: 21711226 Free PMC article. Review.
-
Non-viral chimeric antigen receptor (CAR) T cells going viral.Immunooncol Technol. 2023 Mar 9;18:100375. doi: 10.1016/j.iotech.2023.100375. eCollection 2023 Jun. Immunooncol Technol. 2023. PMID: 37124148 Free PMC article. Review.
References
-
- Hermonat PL, Quirk JG, Bishop BM, Han L. The packaging capacity of adeno-associated virus (AAV) and the potential for wild-type-plus AAV gene therapy vectors. FEBS Lett. 1997;407:78–84. - PubMed
-
- Kumar M, Keller B, Makalou N, Sutton RE. Systematic determination of the packaging limit of lentiviral vectors. Hum Gene Ther. 2001;12:1893–1905. - PubMed
-
- Senior SL, Wade-Martins R. Herpes simplex virus type 1 amplicon vectors for the infectious delivery and expression of genomic DNA loci. Curr Opin Mol Ther. 2005;7:337–345. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
