

Oxidative half-reaction of arabidopsis thaliana sulfite oxidase: generation of superoxide by a peroxisomal enzyme
- PMID: 19875441
- PMCID: PMC2790977
- DOI: 10.1074/jbc.M109.067355
Oxidative half-reaction of arabidopsis thaliana sulfite oxidase: generation of superoxide by a peroxisomal enzyme
Abstract
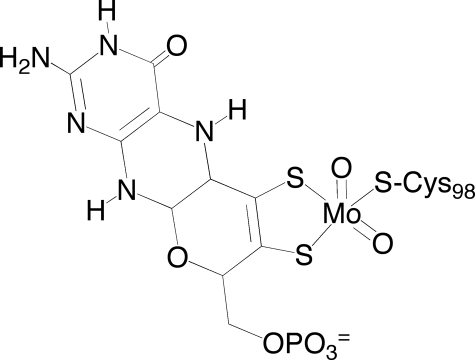
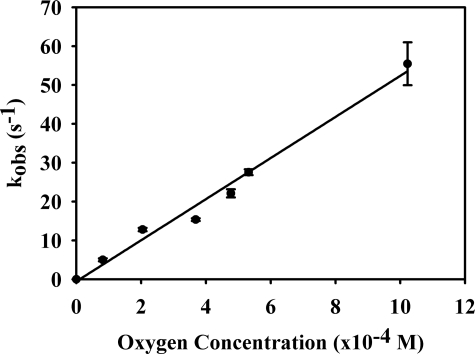
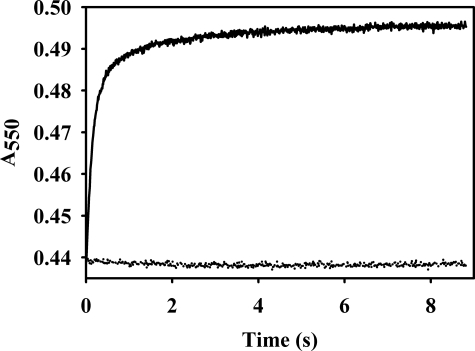
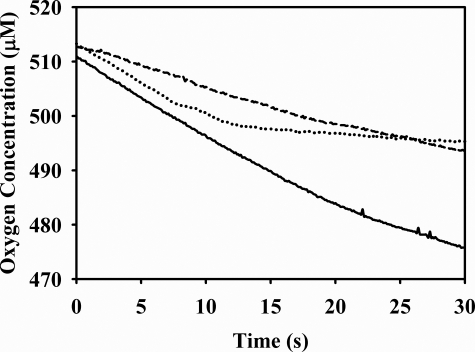
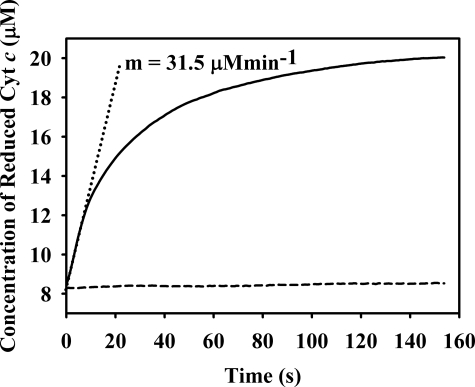
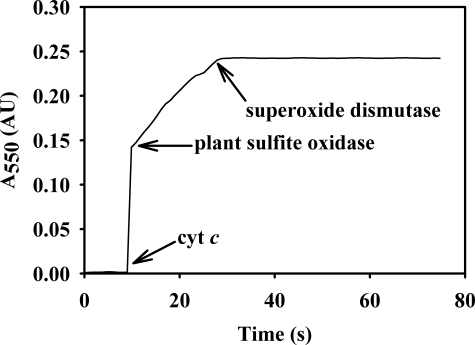
Vertebrate forms of the molybdenum-containing enzyme sulfite oxidase possess a b-type cytochrome prosthetic group that accepts reducing equivalents from the molybdenum center and passes them on to cytochrome c. The plant form of the enzyme, on the other hand, lacks a prosthetic group other than its molybdenum center and utilizes molecular oxygen as the physiological oxidant. Hydrogen peroxide is the ultimate product of the reaction. Here, we present data demonstrating that superoxide is produced essentially quantitatively both in the course of the reaction of reduced enzyme with O(2) and during steady-state turnover and only subsequently decays (presumably noncatalytically) to form hydrogen peroxide. Rapid-reaction kinetic studies directly following the reoxidation of reduced enzyme demonstrate a linear dependence of the rate constant for the reaction on [O(2)] with a second-order rate constant of k(ox) = 8.7 x 10(4) +/- 0.5 x 10(4) m(-1)s(-1). When the reaction is carried out in the presence of cytochrome c to follow superoxide generation, biphasic time courses are observed, indicating that a first equivalent of superoxide is generated in the oxidation of the fully reduced Mo(IV) state of the enzyme to Mo(V), followed by a slower oxidation of the Mo(V) state to Mo(VI). The physiological implications of plant sulfite oxidase as a copious generator of superoxide are discussed.
Figures
References
-
- Cohen H. J., Fridovich I. J. (1971) J. Biol. Chem. 246, 359–366 - PubMed
-
- Niknahad H., O'Brien P. J. (2008) Chem. Biol. Interact. 174, 147–154 - PubMed
-
- Schindelin H., Kisker C., Rajagopalan K. V. (2001) Adv. Protein Chem. 58, 47–94 - PubMed
-
- Hänsch R., Lang C., Rennenberg H., Mendel R. R. (2007) Plant Biol. 9, 589–595 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
