

doi: 10.2337/db09-0580.
Metabolomics applied to diabetes research: moving from information to knowledge
Affiliations
- PMID: 19875619
- PMCID: PMC2768174
- DOI: 10.2337/db09-0580
Item in Clipboard
Metabolomics applied to diabetes research: moving from information to knowledge
Diabetes.
2009 Nov.
No abstract available
Figures

Schematic summary of targeted and nontargeted metabolomics methods. A: When using targeted methods, quantification of specific analytes is facilitated by addition of stable isotope-labeled standards to the sample prior to the sample extraction and derivatization steps. This allows reporting of targeted analytes in true units of measure (e.g., μmol/l). B: In contrast, when performing nontargeted analysis, the goal is usually to obtain a global comparison of a large number of analytes across several classes. This is achieved by assaying replicate samples from contrasting conditions (e.g., drug-treated vs. control cells). Samples are processed and analyzed by MS to generate two independent datasets that are subjected to univariate and multivariate statistical analysis to identify features that are different between the two conditions, although in the absence of added standards, the concentration of the analytes is not reported in true units of measure. In addition, for many of the peaks, the chemical identity of the metabolite cannot be immediately discerned due to limitations in current spectral and chemical standard libraries. PCA, principal components analysis.
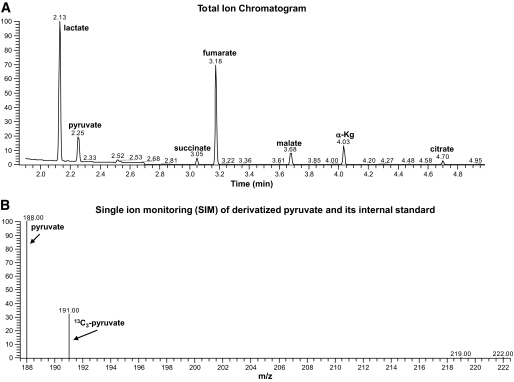
Targeted isotope-dilution analysis of organic acids by GC-MS. When performing targeted isotope-dilution analysis, a group of heavy isotope standards specific to the assay module are added immediately following sample homogenization and prior to chromatographic resolution and MS. A: Total ion chromatogram following GC of a rat liver homogenate in which multiple organic acids are resolved. B: Single ion monitoring for pyruvate and its corresponding internal standard. Each organic acid peak in the chromatogram (A) contains the target analyte and a heavy stable isotope as an internal standard, which are resolved and quantified by mass spectrometry (B) as shown for pyruvate, for example.
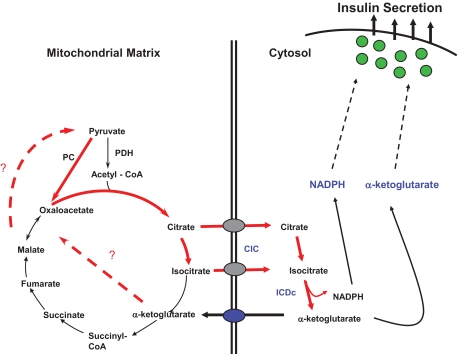
Schematic diagram of a pyruvate/isocitrate cycle implicated in control of GSIS. The cycle is initiated by anaplerotic conversion of pyruvate to oxaloacetate by PC. This leads to accumulation of the TCA cycle intermediates citrate and isocitrate and their export from the mitochondria to the cytosol by the CIC. Citrate is then converted to isocitrate by cytosolic aconitase, and isocitrate is converted to α-ketoglutarate by the cytosolic NADP-dependent ICDc. α-Ketoglutarate can then serve either as a direct signal for insulin secretion, for example, by serving as a substrate for α-ketoglutarate hydroxylases, or be recycled to pyruvate by one of several mitochondrial or cytosolic pathways that remain to be defined (dashed lines). Another byproduct of the pyruvate/isocitrate cycle with potential as an insulin secretagogue is cytosolic NAPDH, possibly acting through Kv channels or the glutathione/glutaredoxin system. This pathway was elucidated by integration of flux analysis by 13C NMR, targeted metabolic profiling by GC-MS and MS-MS, and evaluation of the effects of knockdown of key genes in the pathway, including CIC and ICDc.

Mechanistic models of lipid-induced impairment of muscle insulin action and supporting metabolomics data. Feeding of diets high in fat results in muscle insulin resistance, and recent studies suggest the operation of two possible mechanisms for this effect (A). A prevailing theory is that increased delivery of fat to muscle saturates the capacity for mitochondrial β-oxidation, leading to accumulation of bioactive lipid-derived metabolites such as diacylglycerols and ceramides in the extramitochondrial space and activation of stress/serine kinases that interfere with insulin action. More recent studies have shown that fatty acid oxidation is actually increased in muscle in response to high-fat feeding but with no coordinate increase in TCA cycle activity. This results in accumulation of incompletely oxidized lipids in the mitochondria and depletion of TCA cycle intermediates, possibly resulting in mitochondrial stress and interference with insulin actions. The metabolic changes that underpin this new mechanism were identified by targeted GC-MS of organic acids and MS-MS analysis of acylcarnitines in muscle extracts from lean and obese animals, as summarized in B (data reprinted from ref. with permission). Note that these mechanisms are not mutually exclusive and could work in concert to impair muscle insulin action. CPT1, carnitine palmitoyltransferase 1; ETS, electron transport system; NEFA, nonesterified fatty acid; TCAI, TCA cycle intermediates.

Mechanistic models of lipid-induced impairment of muscle insulin action and supporting metabolomics data. Feeding of diets high in fat results in muscle insulin resistance, and recent studies suggest the operation of two possible mechanisms for this effect (A). A prevailing theory is that increased delivery of fat to muscle saturates the capacity for mitochondrial β-oxidation, leading to accumulation of bioactive lipid-derived metabolites such as diacylglycerols and ceramides in the extramitochondrial space and activation of stress/serine kinases that interfere with insulin action. More recent studies have shown that fatty acid oxidation is actually increased in muscle in response to high-fat feeding but with no coordinate increase in TCA cycle activity. This results in accumulation of incompletely oxidized lipids in the mitochondria and depletion of TCA cycle intermediates, possibly resulting in mitochondrial stress and interference with insulin actions. The metabolic changes that underpin this new mechanism were identified by targeted GC-MS of organic acids and MS-MS analysis of acylcarnitines in muscle extracts from lean and obese animals, as summarized in B (data reprinted from ref. with permission). Note that these mechanisms are not mutually exclusive and could work in concert to impair muscle insulin action. CPT1, carnitine palmitoyltransferase 1; ETS, electron transport system; NEFA, nonesterified fatty acid; TCAI, TCA cycle intermediates.

Metabolite levels are heritable and can be mapped to specific chromosomes in mice. In this particular study, diabetes-resistant B6-ob/ob mice were bred with diabetes-susceptible BTBR-ob/ob mice to create a cohort of F2 mice in which individual mice had a wide variation in blood glucose levels. Whole-genome SNP analysis was integrated with microarray and targeted GC-MS– and MS-MS–based analysis of metabolites of liver extracts from the F2 animals. This analysis revealed that metabolites can be mapped to specific chromosomes. Each row represents a SNP marker, and each column represents a metabolite. The LOD color scale is indicated, showing blue when the B6 allele at that marker results in an elevated level of the metabolite and red/yellow when the BTBR allele is dominant. Note the clusters of amino acids that map in common to regions on chromosomes 8 and 9, suggesting the presence of a gene that controls their levels. Figure reproduced with permission from ref. . LOD, logarithm of odds.

A novel metabolic regulatory network predicted by integration of genomic, transcriptomic, and metabolomic profiling. Whole-genome mapping was integrated with transcriptomic and metabolomic analysis of liver samples in F2 mice from a cross of diabetes-resistant B6-ob/ob mice and diabetes-susceptible BTBR-ob/ob mice. Networks of transcripts (grey ovals) and metabolites (rectangle) were identified by computational analysis (63). The network shown here predicts that the amino acid glutamate/glutamine (Glx) regulates expression of the key gluconeogenic enzyme phosphoenolpyruvate carboxykinase (Pck1) via regulation of alanine/glyoxylate aminotransferase (Agxt) and arginase1 (Arg1). Consistent with this prediction, glutamine addition to mouse hepatocytes strongly induces expression of Agxt, Arg1, and Pck1 (20). Figure reproduced with permission from ref. .

A branched-chain amino acid–related metabolite cluster that correlates with insulin resistance in humans. Targeted metabolic profiling by MS-MS and GC-MS was performed on obese, insulin-resistant, but nondiabetic humans and a group of lean controls. Principal components analysis (PCA) revealed that the cluster of statistically related metabolites (principal component) with the strongest differences between obese and lean subjects was one comprised of the branched-chain amino acids leucine/isoleucine and valine, glutamate/glutamine (glx), C3 and C5 acylcarnitines, and the aromatic amino acids phenylalanine and tyrosine. A plot of each individual's principal component score against their homeostasis model assesment index score (a measure of insulin sensitivity) is shown, indicating significant correlation (r = 0.58; P < 0.0001). Data reprinted with permission from ref. .
References
-
- Ridderstråle M, Groop L: Genetic dissection of type 2 diabetes. Mol Cell Endocrinol 2009;297:10–17 - PubMed
-
- Muoio DM, Newgard CB: Molecular and metabolic mechanisms of insulin resistance and β-cell failure in type 2 diabetes. Nat Rev Mol Cell Biol 2008;9:193–205 - PubMed
-
- Human Metabolome Project: Human Metabolome Database-Canada [website]. Available at http://www.hmdb.ca/ Accessed 19 September 2009
-
- Castle AL, Fiehn O, Kaddurah-Daouk R, Lindon JC: Metabolomics Standards Workshop and the development of international standards for reporting metabolomics experimental results. Brief Bioinform 2006;7:159–165 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
