

A gastrin precursor, gastrin-gly, upregulates VEGF expression in colonic epithelial cells through an HIF-1-independent mechanism
- PMID: 19876923
- PMCID: PMC3682422
- DOI: 10.1002/ijc.25001
A gastrin precursor, gastrin-gly, upregulates VEGF expression in colonic epithelial cells through an HIF-1-independent mechanism
Abstract
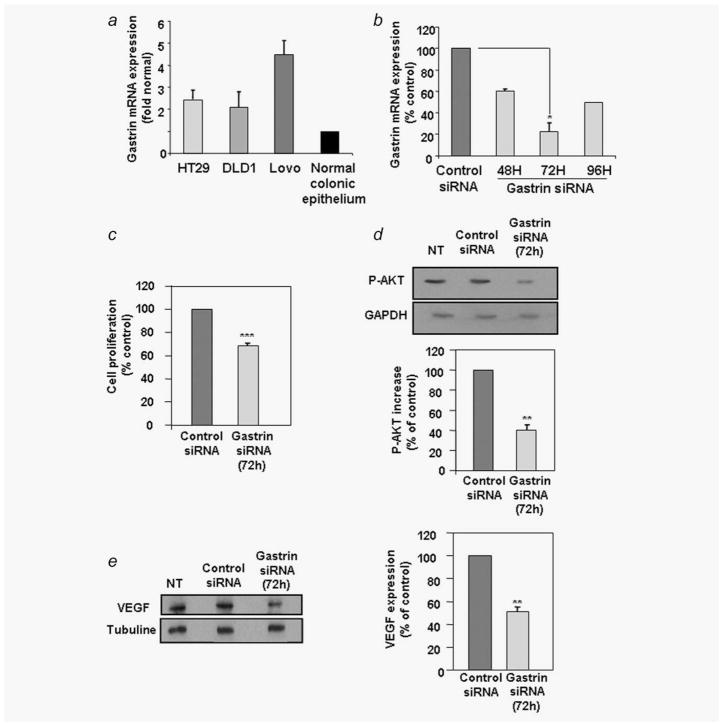
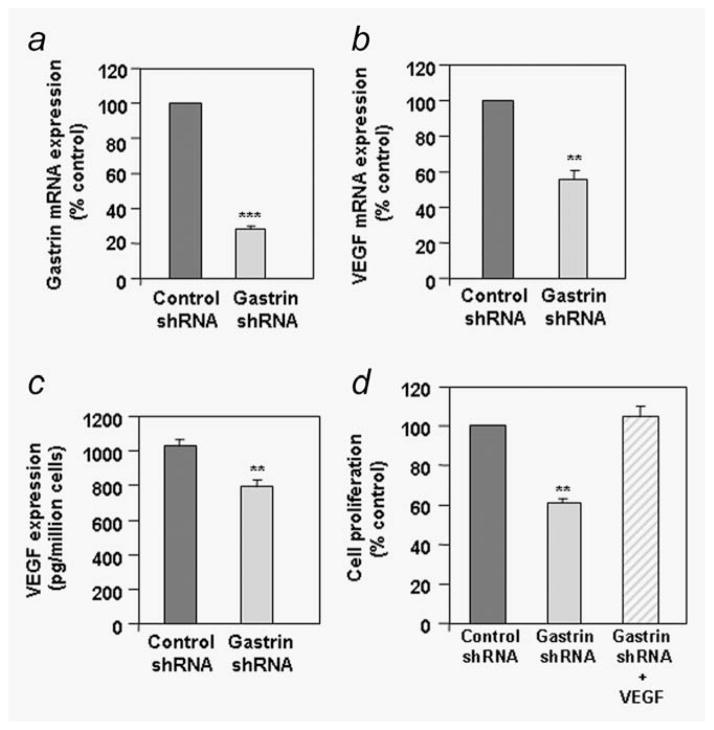
One of the major angiogenic factor released by tumor cells is VEGF. Its high expression is correlated with poor prognosis in colorectal tumors. In colon cancer, gastrin gene expression is also upregulated. In these tumors, gastrin precursors are mainly produced and act as growth factors. Recently, a study has also shown that the gastrin precursor, G-gly induced in vitro tubules formation by vascular endothelial cells suggesting a potential proangiogenic role. Here, we demonstrate that stimulation of human colorectal cancer cell lines with G-gly increases the expression of the proangiogenic factor VEGF at the mRNA and protein levels. In addition, blocking the progastrin autocrine loop leads to a downregulation of VEGF. Although HIF-1 is a major transcriptional activator for VEGF our results suggest an alternative mechanism for VEGF regulation in normoxic conditions, independent of HIF-1 that involves the PI3K/AKT pathway. Indeed we show that G-gly does not lead to HIF-1 accumulation in colon cancer cells. Moreover, we found that G-gly activates the PI3K/AKT pathway and inhibition of this pathway reverses the effects of G-gly observed on VEGF mRNA and protein levels. In correlation with these results, we observed in vivo, on colon tissue sections from transgenic mice overexpressing G-gly, an increase in VEGF expression in absence of HIF-1 accumulation. In conclusion, our study demonstrates that gastrin precursors, known to promote colon epithelial cells proliferation and survival can also contribute to the angiogenesis process by stimulating the expression of the proangiogenic factor VEGF via the PI3K pathway and independently of hypoxia conditions.
Figures





Similar articles
-
Celecoxib Down-Regulates the Hypoxia-Induced Expression of HIF-1α and VEGF Through the PI3K/AKT Pathway in Retinal Pigment Epithelial Cells.Cell Physiol Biochem. 2017;44(4):1640-1650. doi: 10.1159/000485764. Epub 2017 Dec 6. Cell Physiol Biochem. 2017. PMID: 29216640
-
Role of PI3K/Akt and MEK/ERK in mediating hypoxia-induced expression of HIF-1alpha and VEGF in laser-induced rat choroidal neovascularization.Invest Ophthalmol Vis Sci. 2009 Apr;50(4):1873-9. doi: 10.1167/iovs.08-2591. Epub 2008 Dec 20. Invest Ophthalmol Vis Sci. 2009. PMID: 19098317
-
MMP-2 alters VEGF expression via alphaVbeta3 integrin-mediated PI3K/AKT signaling in A549 lung cancer cells.Int J Cancer. 2010 Sep 1;127(5):1081-95. doi: 10.1002/ijc.25134. Int J Cancer. 2010. PMID: 20027628 Free PMC article.
-
Delphinidin inhibits angiogenesis through the suppression of HIF-1α and VEGF expression in A549 lung cancer cells.Oncol Rep. 2017 Feb;37(2):777-784. doi: 10.3892/or.2016.5296. Epub 2016 Dec 7. Oncol Rep. 2017. PMID: 27959445
-
Role of Annexin-II in GI cancers: interaction with gastrins/progastrins.Cancer Lett. 2007 Jul 8;252(1):19-35. doi: 10.1016/j.canlet.2006.11.012. Epub 2006 Dec 22. Cancer Lett. 2007. PMID: 17188424 Free PMC article. Review.
Cited by
-
Association of helicobacter pylori infection and colon cancer.J Clin Med Res. 2012 Jun;4(3):172-6. doi: 10.4021/jocmr880w. Epub 2012 May 15. J Clin Med Res. 2012. PMID: 22719803 Free PMC article.
-
Progastrin a new pro-angiogenic factor in colorectal cancer.Oncogene. 2015 Jun 11;34(24):3120-30. doi: 10.1038/onc.2014.255. Epub 2014 Aug 11. Oncogene. 2015. PMID: 25109333
-
Progastrin: a potential predictive marker of liver metastasis in colorectal cancer.Int J Colorectal Dis. 2017 Jul;32(7):1061-1064. doi: 10.1007/s00384-017-2822-8. Epub 2017 Apr 21. Int J Colorectal Dis. 2017. PMID: 28432443
-
Progastrin Peptides Increase the Risk of Developing Colonic Tumors: Impact on Colonic Stem Cells.Curr Colorectal Cancer Rep. 2012 Dec;8(4):277-289. doi: 10.1007/s11888-012-0144-3. Curr Colorectal Cancer Rep. 2012. PMID: 23226720 Free PMC article.
-
Induction of gastrin expression in gastrointestinal cells by hypoxia or cobalt is independent of hypoxia-inducible factor (HIF).Endocrinology. 2012 Jul;153(7):3006-16. doi: 10.1210/en.2011-2069. Epub 2012 May 16. Endocrinology. 2012. PMID: 22593272 Free PMC article.
References
-
- Guba M, Seeliger H, Kleespies A, Jauch KW, Bruns C. Vascular endothelial growth factor in colorectal cancer. Int J Colorectal Dis. 2004;19:510–7. - PubMed
-
- Dufresne M, Seva C, Fourmy D. Cholecystokinin and gastrin receptors. Physiol Rev. 2006;86:805–47. - PubMed
-
- Lei S, Dubeykovskiy A, Chakladar A, Wojtukiewicz L, Wang TC. The murine gastrin promoter is synergistically activated by transforming growth factor-beta/Smad and Wnt signaling pathways. J Biol Chem. 2004;279:42492–502. - PubMed
-
- Chakladar A, Dubeykovskiy A, Wojtukiewicz LJ, Pratap J, Lei S, Wang TC. Synergistic activation of the murine gastrin promoter by oncogenic Ras and beta-catenin involves SMAD recruitment. Biochem Biophys Res Commun. 2005;336:190–6. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
