

Mst1 and Mst2 maintain hepatocyte quiescence and suppress hepatocellular carcinoma development through inactivation of the Yap1 oncogene
- PMID: 19878874
- PMCID: PMC3023165
- DOI: 10.1016/j.ccr.2009.09.026
Mst1 and Mst2 maintain hepatocyte quiescence and suppress hepatocellular carcinoma development through inactivation of the Yap1 oncogene
Abstract
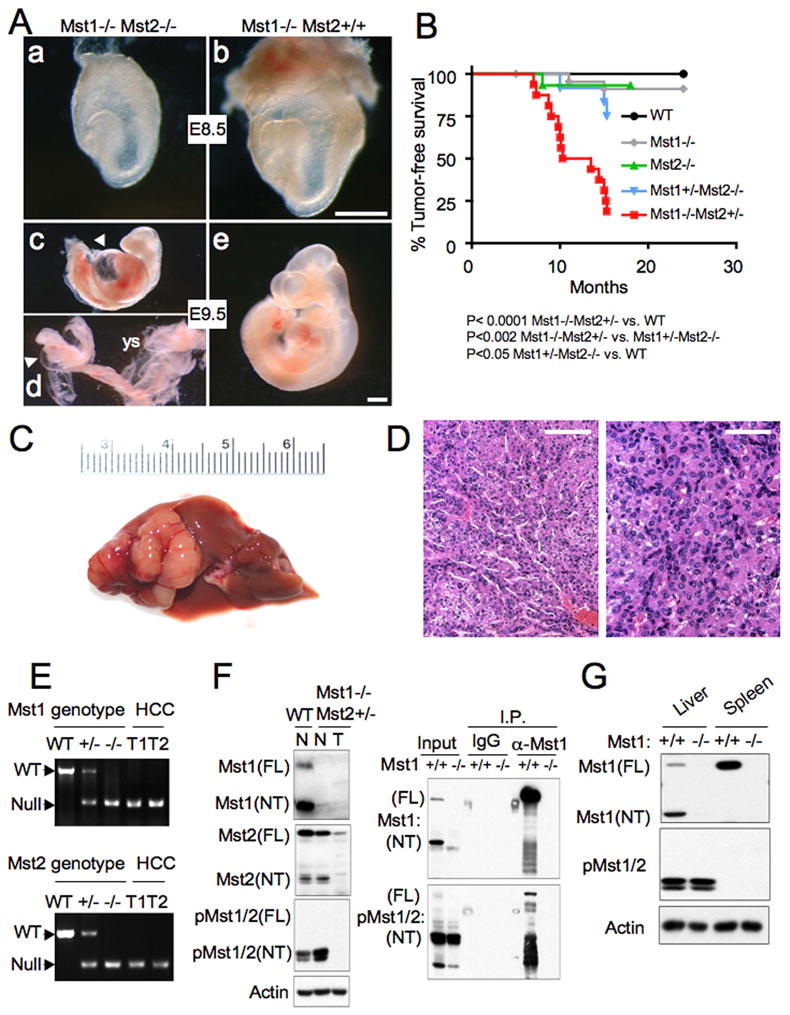
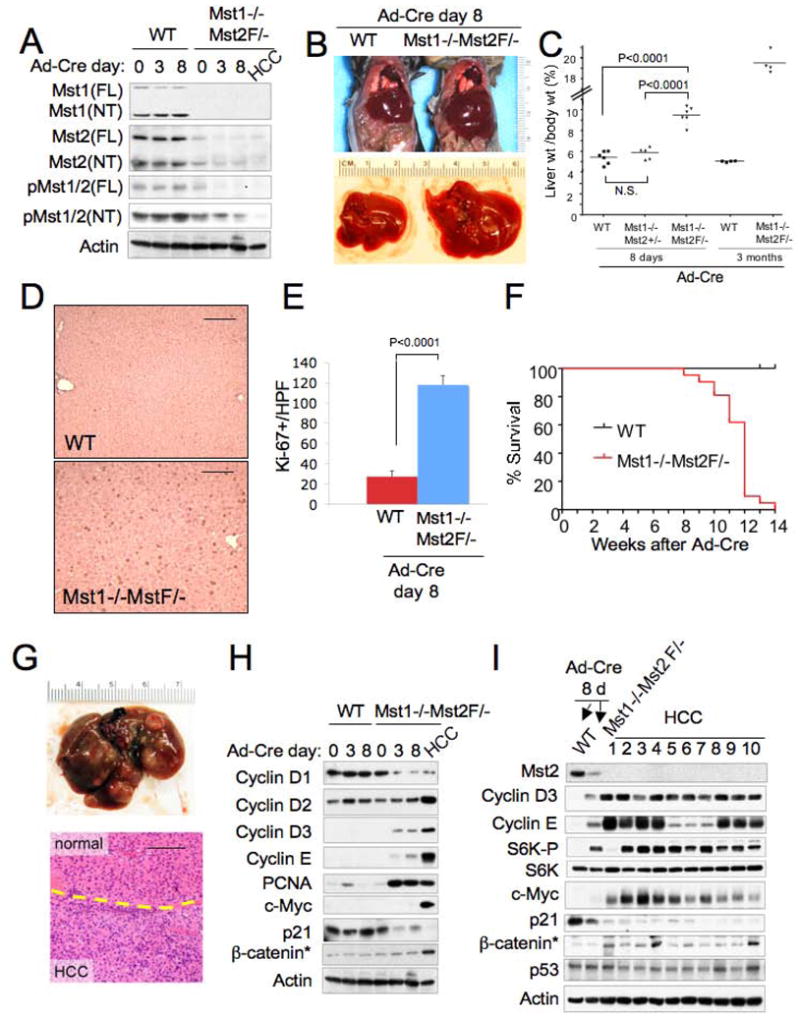
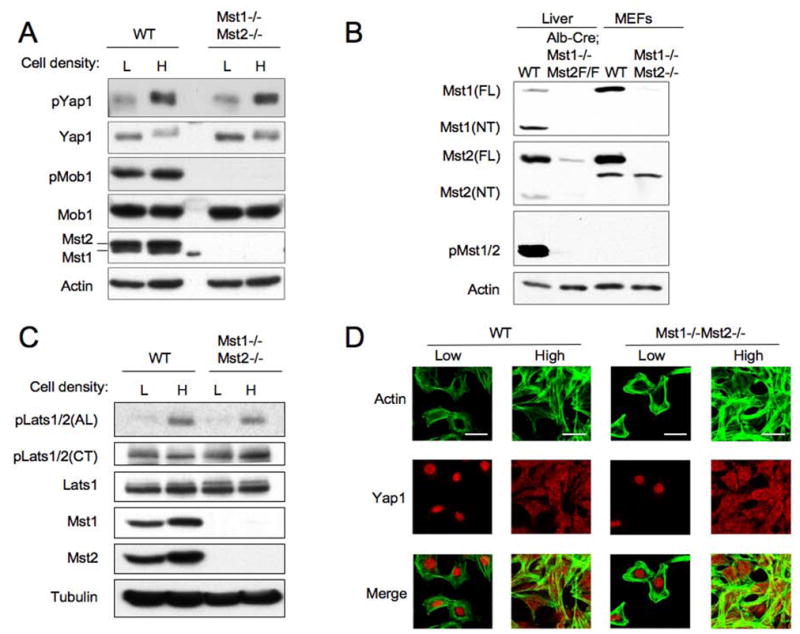
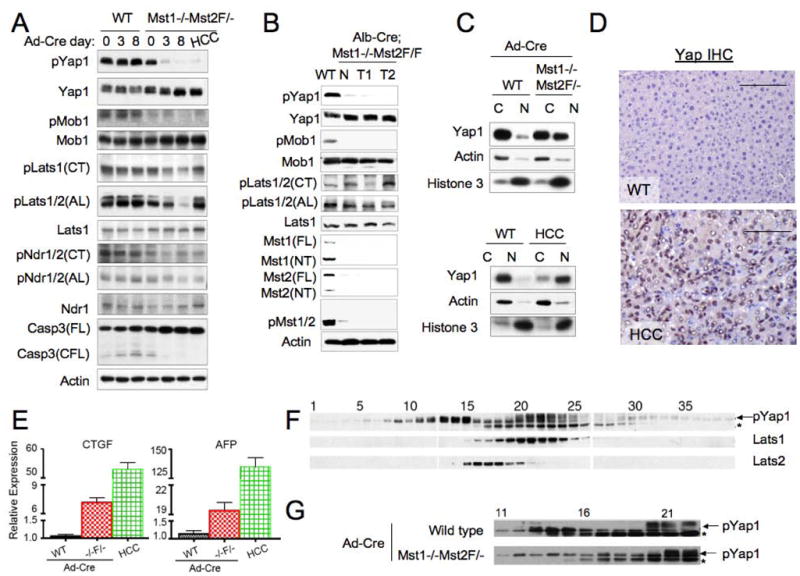
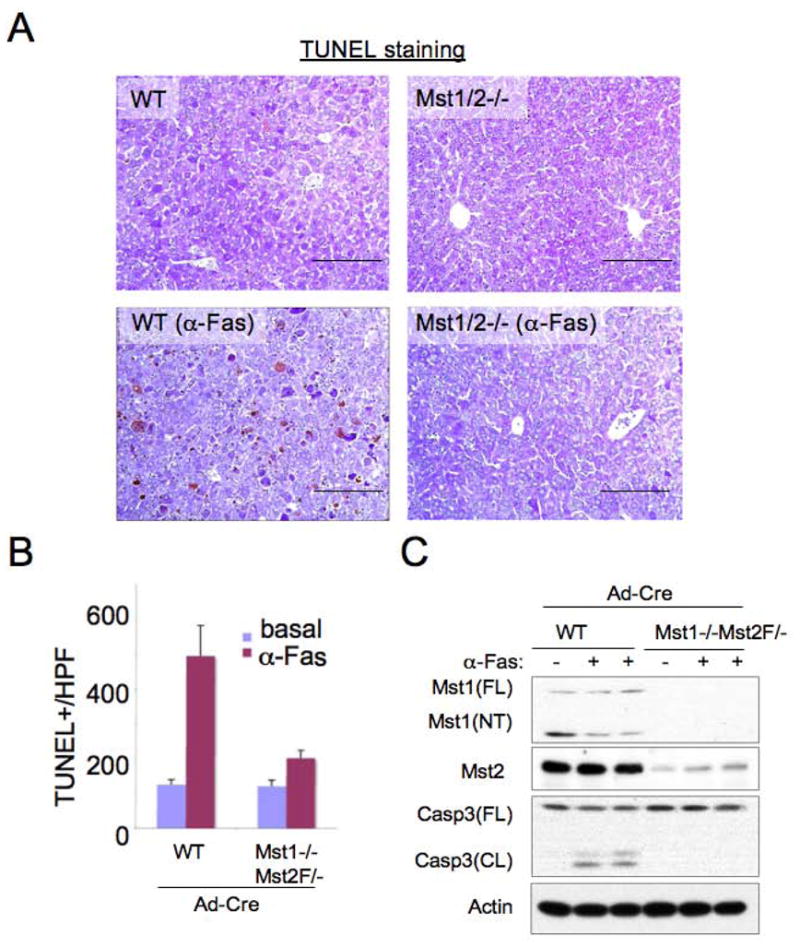
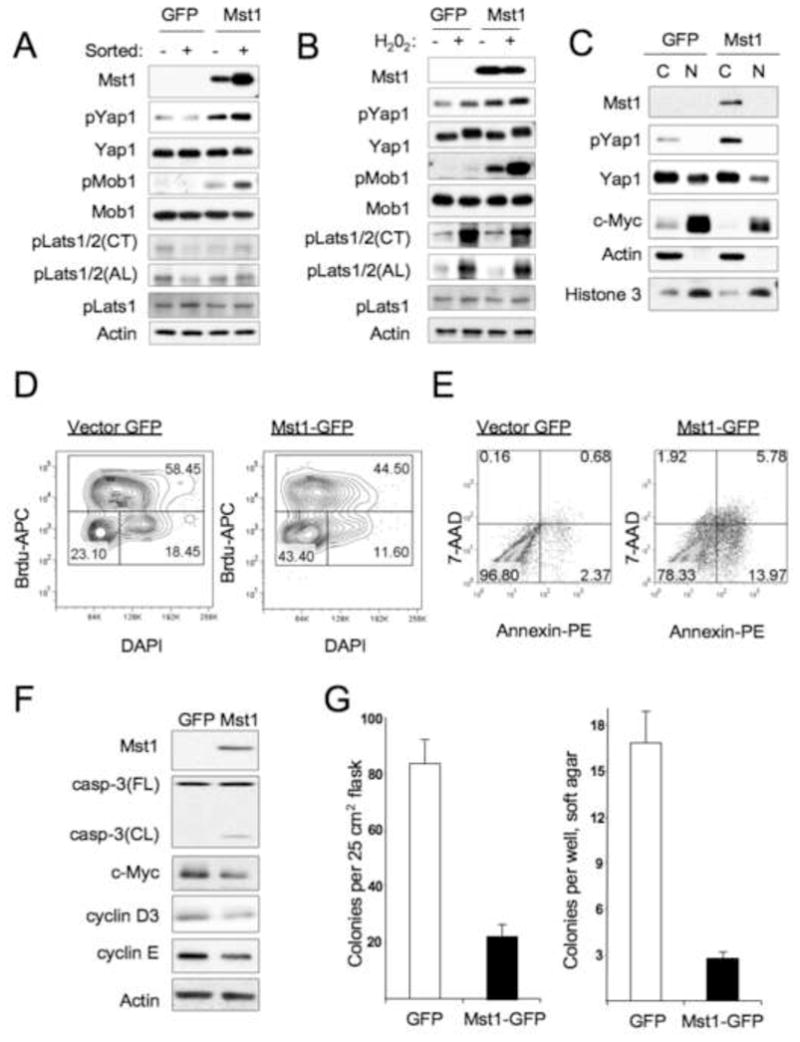
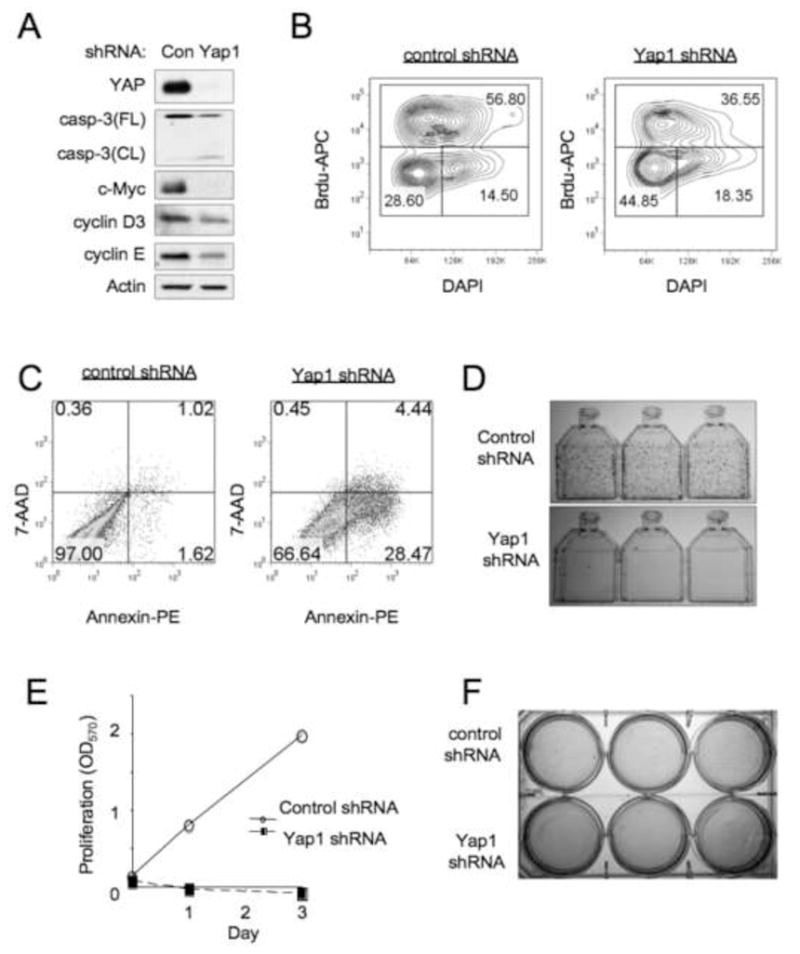
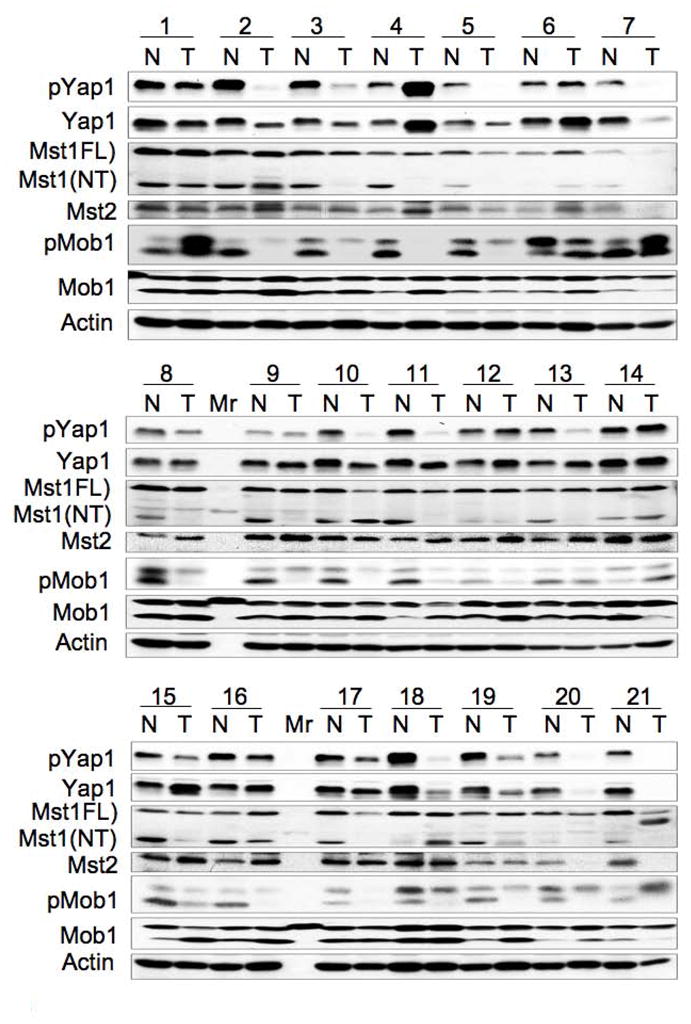
Hippo-Lats-Yorkie signaling regulates tissue overgrowth and tumorigenesis in Drosophila. We show that the Mst1 and Mst2 protein kinases, the mammalian Hippo orthologs, are cleaved and constitutively activated in the mouse liver. Combined Mst1/2 deficiency in the liver results in loss of inhibitory Ser127 phosphorylation of the Yorkie ortholog, Yap1, massive overgrowth, and hepatocellular carcinoma (HCC). Reexpression of Mst1 in HCC-derived cell lines promotes Yap1 Ser127 phosphorylation and inactivation and abrogates their tumorigenicity. Notably, Mst1/2 inactivates Yap1 in liver through an intermediary kinase distinct from Lats1/2. Approximately 30% of human HCCs show low Yap1(Ser127) phosphorylation and a majority exhibit loss of cleaved, activated Mst1. Mst1/2 inhibition of Yap1 is an important pathway for tumor suppression in liver relevant to human HCC.
Figures
Comment in
-
Mst out and HCC in.Cancer Cell. 2009 Nov 6;16(5):363-4. doi: 10.1016/j.ccr.2009.10.008. Cancer Cell. 2009. PMID: 19878866
-
Hippo tumor supressor pathway: novel implications for the treatment of hepatocellular carcinoma.Gastroenterology. 2010 Aug;139(2):692-4. doi: 10.1053/j.gastro.2010.06.036. Epub 2010 Jun 25. Gastroenterology. 2010. PMID: 20600057 Free PMC article. No abstract available.
References
-
- Calvisi DF, Ladu S, Gorden A, Farina M, Conner EA, Lee JS, Factor VM, Thorgeirsson SS. Ubiquitous activation of Ras and Jak/Stat pathways in human HCC. Gastroenterology. 2006;130:1117–1128. - PubMed
-
- Camargo FD, Gokhale S, Johnnidis JB, Fu D, Bell GW, Jaenisch R, Brummelkamp TR. YAP1 increases organ size and expands undifferentiated progenitor cells. Curr Biol. 2007;17:2054–2060. - PubMed
-
- Chan EH, Nousiainen M, Chalamalasetty RB, Schafer A, Nigg EA, Sillje HH. The Ste20-like kinase Mst2 activates the human large tumor suppressor kinase Lats1. Oncogene. 2005;24:2076–2086. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
