

Dissection of Rad9 BRCT domain function in the mitotic checkpoint response to telomere uncapping
- PMID: 19880356
- PMCID: PMC2787662
- DOI: 10.1016/j.dnarep.2009.09.010
Dissection of Rad9 BRCT domain function in the mitotic checkpoint response to telomere uncapping
Abstract
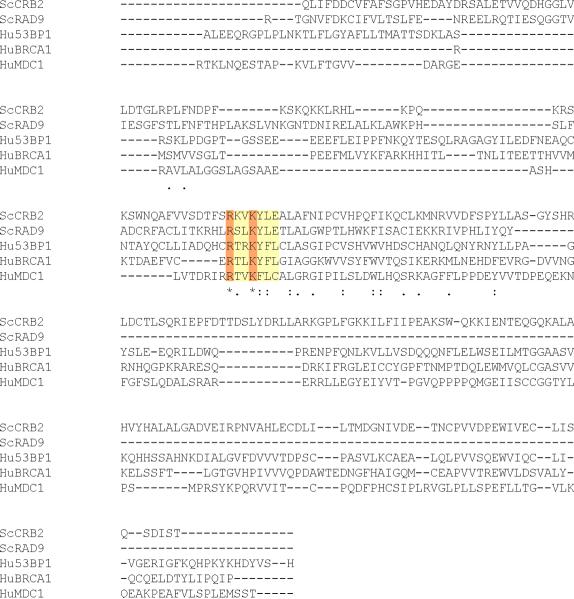
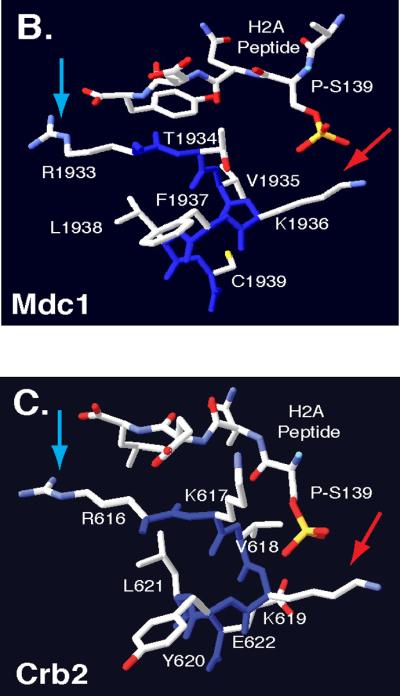
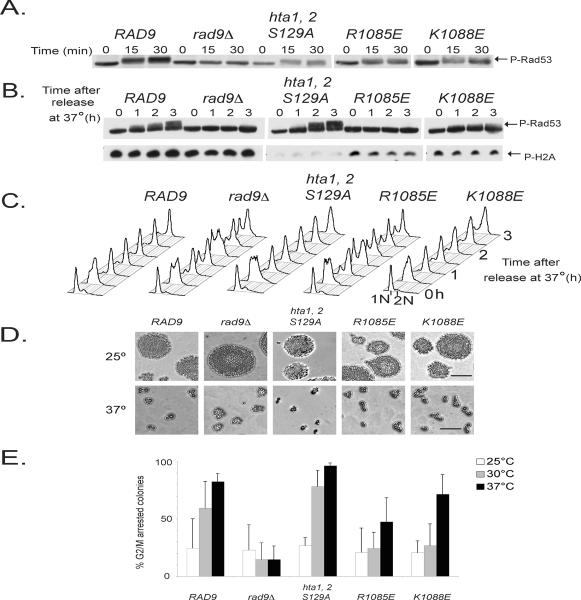
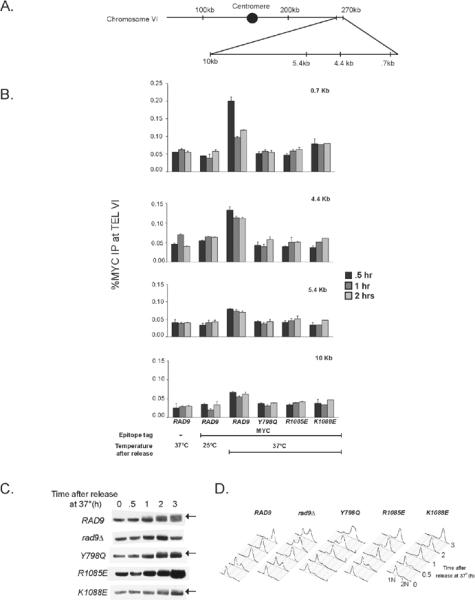
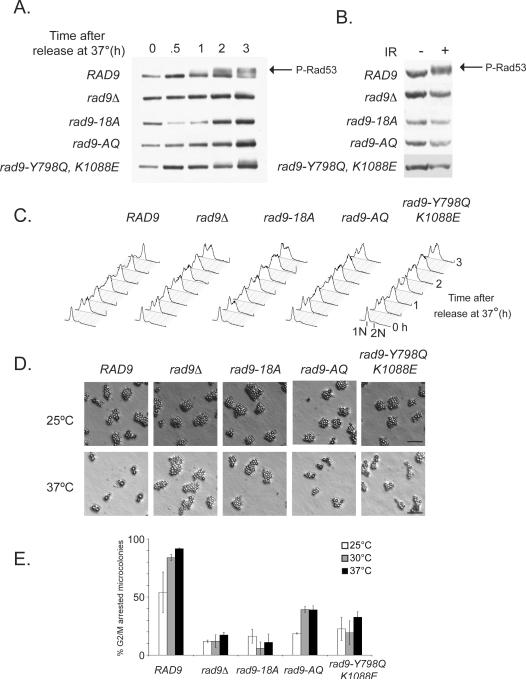
In Saccharomyces cerevisiae, destabilizing telomeres, via inactivation of telomeric repeat binding factor Cdc13, induces a cell cycle checkpoint that arrests cells at the metaphase to anaphase transition--much like the response to an unrepaired DNA double strand break (DSB). Throughout the cell cycle, the multi-domain adaptor protein Rad9 is required for the activation of checkpoint effector kinase Rad53 in response to DSBs and is similarly necessary for checkpoint signaling in response to telomere uncapping. Rad53 activation in G1 and S phase depends on Rad9 association with modified chromatin adjacent to DSBs, which is mediated by Tudor domains binding histone H3 di-methylated at K79 and BRCT domains to histone H2A phosphorylated at S129. Nonetheless, Rad9 Tudor or BRCT mutants can initiate a checkpoint response to DNA damage in nocodazole-treated cells. Mutations affecting di-methylation of H3 K79, or its recognition by Rad9 enhance 5' strand resection upon telomere uncapping, and potentially implicate Rad9 chromatin binding in the checkpoint response to telomere uncapping. Indeed, we report that Rad9 binds to sub-telomeric chromatin, upon telomere uncapping, up to 10 kb from the telomere. Rad9 binding occurred within 30 min after inactivating Cdc13, preceding Rad53 phosphorylation. In turn, Rad9 Tudor and BRCT domain mutations blocked chromatin binding and led to attenuated checkpoint signaling as evidenced by decreased Rad53 phosphorylation and impaired cell cycle arrest. Our work identifies a role for Rad9 chromatin association, during mitosis, in the DNA damage checkpoint response to telomere uncapping, suggesting that chromatin binding may be an initiating event for checkpoints throughout the cell cycle.
Figures
Similar articles
-
Yeast G1 DNA damage checkpoint regulation by H2A phosphorylation is independent of chromatin remodeling.Proc Natl Acad Sci U S A. 2006 Sep 12;103(37):13771-6. doi: 10.1073/pnas.0511192103. Epub 2006 Aug 29. Proc Natl Acad Sci U S A. 2006. PMID: 16940359 Free PMC article.
-
Dynamics of Rad9 chromatin binding and checkpoint function are mediated by its dimerization and are cell cycle-regulated by CDK1 activity.PLoS Genet. 2010 Aug 5;6(8):e1001047. doi: 10.1371/journal.pgen.1001047. PLoS Genet. 2010. PMID: 20700441 Free PMC article.
-
Histone methyltransferase Dot1 and Rad9 inhibit single-stranded DNA accumulation at DSBs and uncapped telomeres.EMBO J. 2008 May 21;27(10):1502-12. doi: 10.1038/emboj.2008.81. Epub 2008 Apr 17. EMBO J. 2008. PMID: 18418382 Free PMC article.
-
Role of the Saccharomyces cerevisiae Rad9 protein in sensing and responding to DNA damage.Biochem Soc Trans. 2003 Feb;31(Pt 1):242-6. doi: 10.1042/bst0310242. Biochem Soc Trans. 2003. PMID: 12546694 Review.
-
Contributions of Rad9 to tumorigenesis.J Cell Biochem. 2012 Mar;113(3):742-51. doi: 10.1002/jcb.23424. J Cell Biochem. 2012. PMID: 22034047 Free PMC article. Review.
Cited by
-
Rad9 interacts with Aft1 to facilitate genome surveillance in fragile genomic sites under non-DNA damage-inducing conditions in S. cerevisiae.Nucleic Acids Res. 2014 Nov 10;42(20):12650-67. doi: 10.1093/nar/gku915. Epub 2014 Oct 9. Nucleic Acids Res. 2014. PMID: 25300486 Free PMC article.
-
BRCT domain interactions with phospho-histone H2A target Crb2 to chromatin at double-strand breaks and maintain the DNA damage checkpoint.Mol Cell Biol. 2010 Oct;30(19):4732-43. doi: 10.1128/MCB.00413-10. Epub 2010 Aug 2. Mol Cell Biol. 2010. PMID: 20679485 Free PMC article.
-
The many faces of histone H3K79 methylation.Mutat Res Rev Mutat Res. 2016 Apr-Jun;768:46-52. doi: 10.1016/j.mrrev.2016.03.005. Epub 2016 Mar 31. Mutat Res Rev Mutat Res. 2016. PMID: 27234562 Free PMC article. Review.
-
DNA Damage-Induced Phosphorylation of Histone H2A at Serine 15 Is Linked to DNA End Resection.Mol Cell Biol. 2021 Nov 22;41(12):e0005621. doi: 10.1128/MCB.00056-21. Epub 2021 Sep 27. Mol Cell Biol. 2021. PMID: 34570618 Free PMC article.
-
Epigenetic modifications in double-strand break DNA damage signaling and repair.Clin Cancer Res. 2010 Sep 15;16(18):4543-52. doi: 10.1158/1078-0432.CCR-10-0513. Epub 2010 Sep 7. Clin Cancer Res. 2010. PMID: 20823147 Free PMC article. Review.
References
-
- Harrison JC, Haber JE. Surviving the breakup: the DNA damage checkpoint. Annu Rev Genet. 2006;40:209–235. - PubMed
-
- Melo J, Toczyski D. A unified view of the DNA-damage checkpoint. Curr Opin Cell Biol. 2002;14:237–245. - PubMed
-
- McGowan CH, Russell P. The DNA damage response: sensing and signaling. Curr Opin Cell Biol. 2004;16:629–633. - PubMed
-
- Lisby M, Barlow JH, Burgess RC, Rothstein R. Choreography of the DNA damage response: spatiotemporal relationships among checkpoint and repair proteins. Cell. 2004;118:699–713. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
