

Spermatogonial stem cells in higher primates: are there differences from those in rodents?
- PMID: 19880674
- PMCID: PMC2895987
- DOI: 10.1530/REP-09-0255
Spermatogonial stem cells in higher primates: are there differences from those in rodents?
Abstract
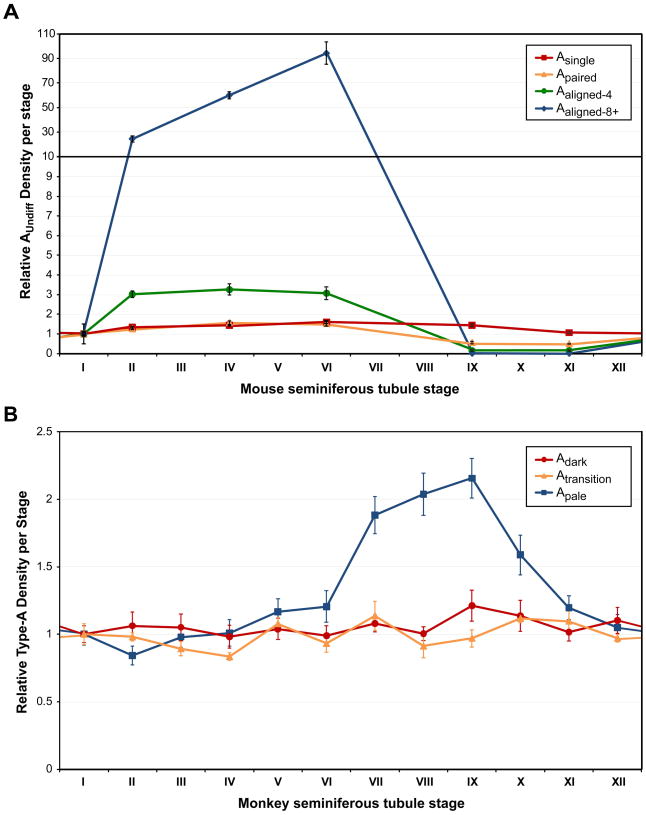
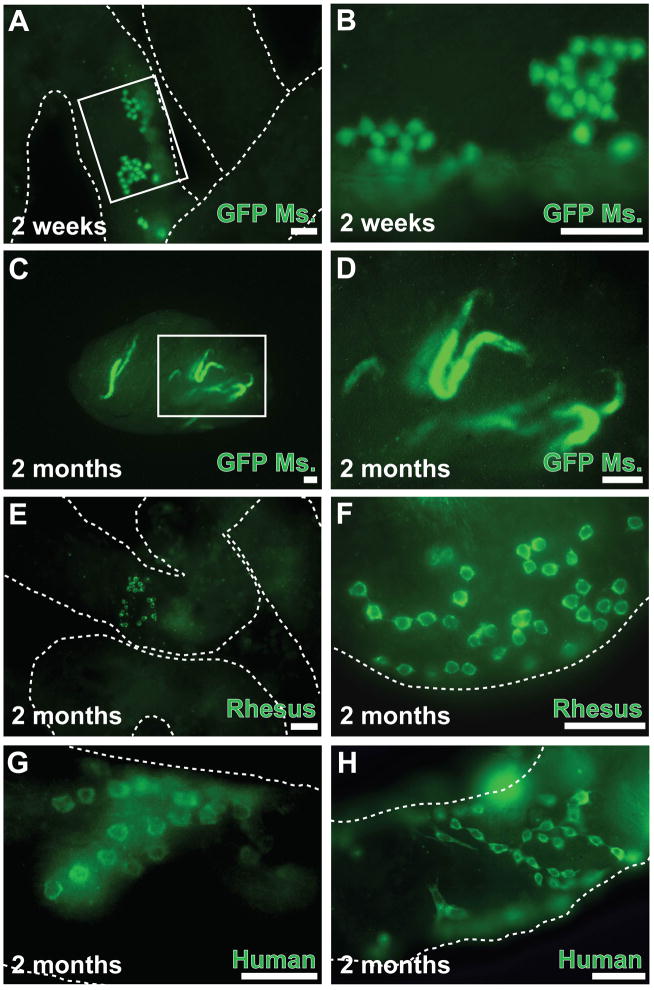
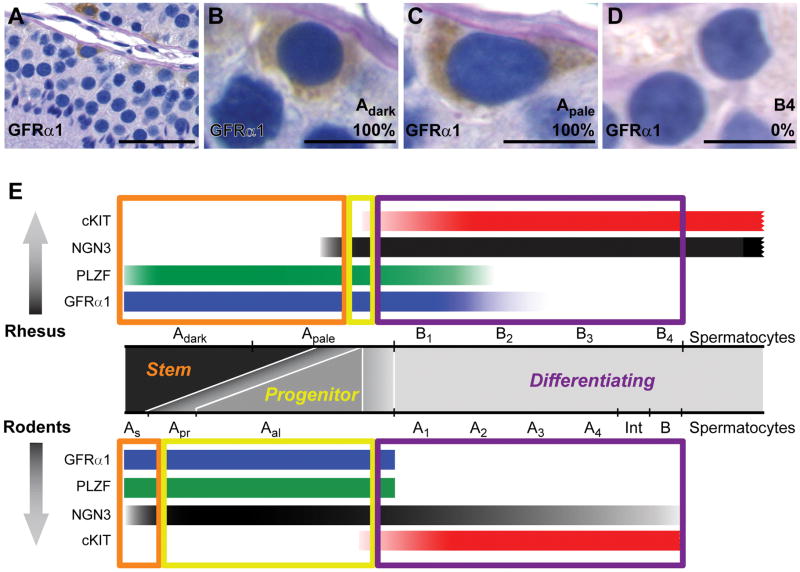
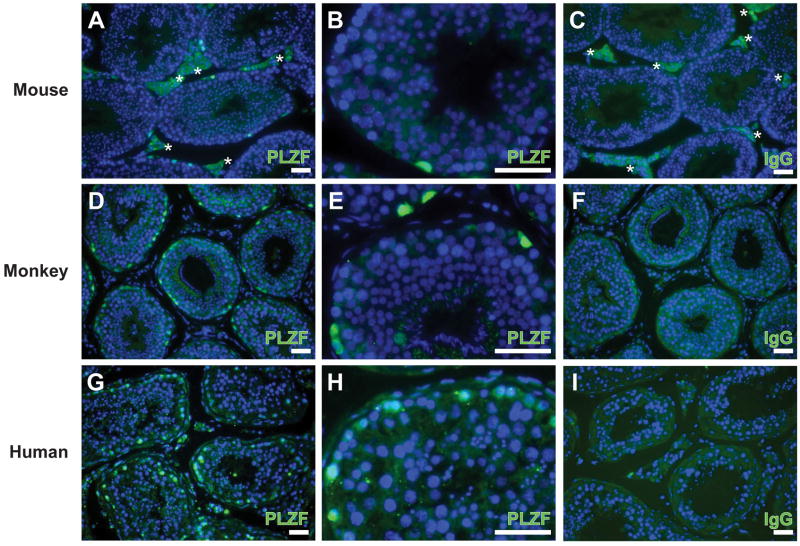
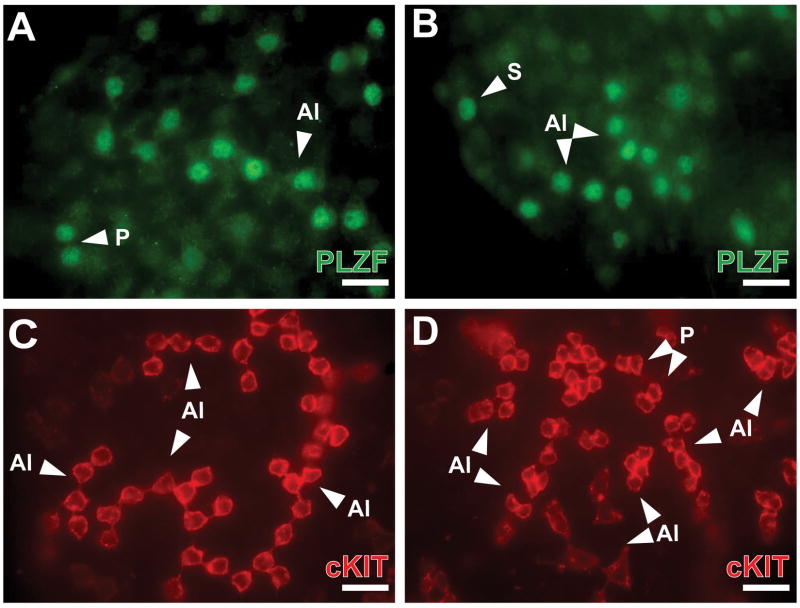
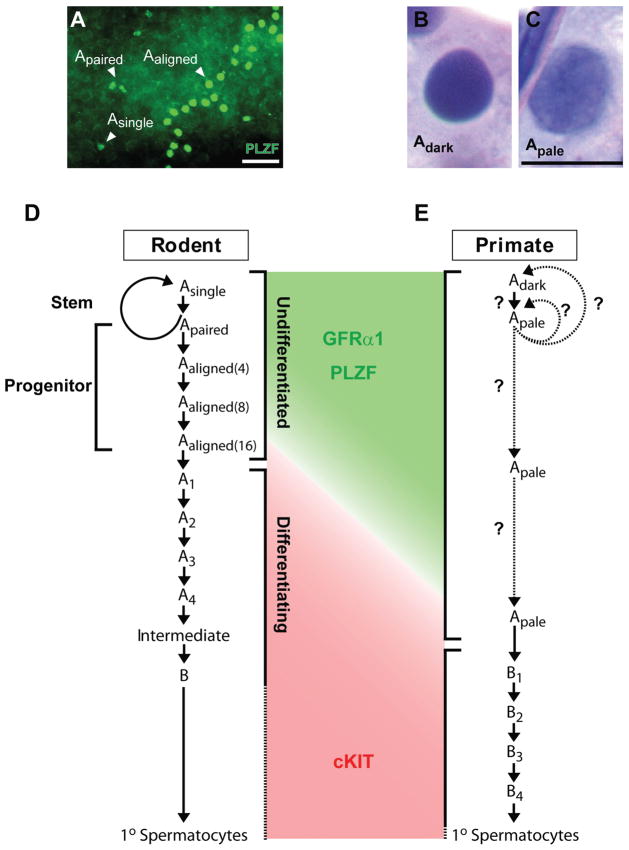
Spermatogonial stem cells (SSCs) maintain spermatogenesis throughout the reproductive life of mammals. While A(single) spermatogonia comprise the rodent SSC pool, the identity of the stem cell pool in the primate spermatogenic lineage is not well established. The prevailing model is that primate spermatogenesis arises from A(dark) and A(pale) spermatogonia, which are considered to represent reserve and active stem cells respectively. However, there is limited information about how the A(dark) and A(pale) descriptions of nuclear morphology correlate with the clonal (A(single), A(paired), and A(aligned)), molecular (e.g. GFRalpha1 (GFRA1) and PLZF), and functional (SSC transplantation) descriptions of rodent SSCs. Thus, there is a need to investigate primate SSCs using criteria, tools, and approaches that have been used to investigate rodent SSCs over the past two decades. SSCs have potential clinical application for treating some cases of male infertility, providing impetus for characterizing and learning to manipulate these adult tissue stem cells in primates (nonhuman and human). This review recounts the development of a xenotransplant assay for functional identification of primate SSCs and progress dissecting the molecular and clonal characteristics of the primate spermatogenic lineage. These observations highlight the similarities and potential differences between rodents and primates regarding the SSC pool and the kinetics of spermatogonial self-renewal and clonal expansion. With new tools and reagents for studying primate spermatogonia, the field is poised to develop and test new hypotheses about the biology and regenerative capacity of primate SSCs.
Conflict of interest statement
Figures
References
-
- Amann RP. The cycle of the seminiferous epithelium in humans: a need to revisit? J Androl. 2008;29:469–487. - PubMed
-
- Barroca V, Lassalle B, Coureuil M, Louis JP, Le PF, Testart J, Allemand I, Riou L, Fouchet P. Mouse differentiating spermatogonia can generate germinal stem cells in vivo. Nat Cell Biol. 2009;11:190–196. - PubMed
-
- Bartmanska J, Clermont Y. Renewal of type A spermatogonia in adult rats. Cell Tissue Kinet. 1983;16:135–143. - PubMed
-
- Brinster CJ, Ryu BY, Avarbock MR, Karagenc L, Brinster RL, Orwig KE. Restoration of Fertility by Germ Cell Transplantation Requires Effective Recipient Preparation. Biology of Reproduction. 2003;69:412–420. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
