

Role of mitogen-activated protein kinase Sty1 in regulation of eukaryotic initiation factor 2alpha kinases in response to environmental stress in Schizosaccharomyces pombe
- PMID: 19880757
- PMCID: PMC2805304
- DOI: 10.1128/EC.00185-09
Role of mitogen-activated protein kinase Sty1 in regulation of eukaryotic initiation factor 2alpha kinases in response to environmental stress in Schizosaccharomyces pombe
Abstract
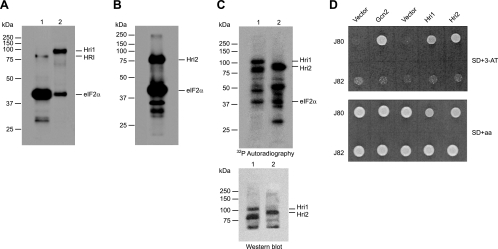
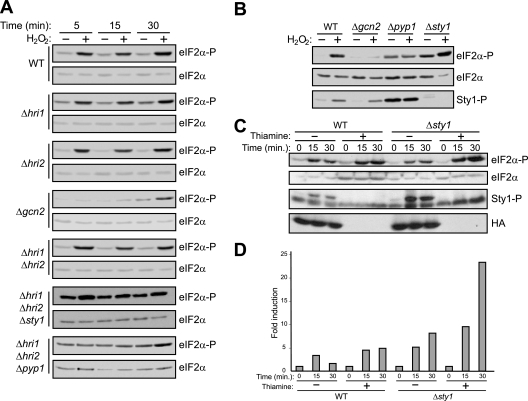
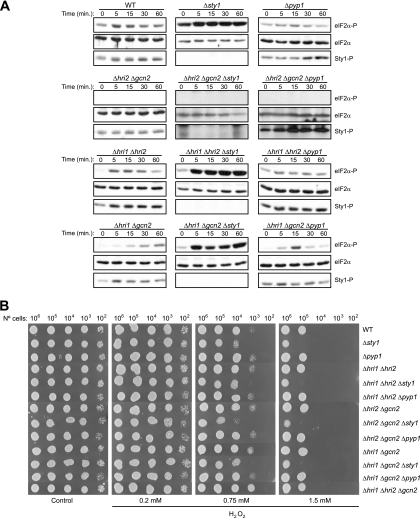
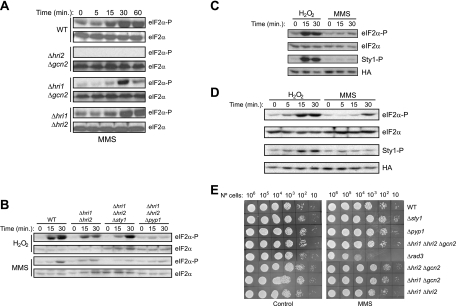
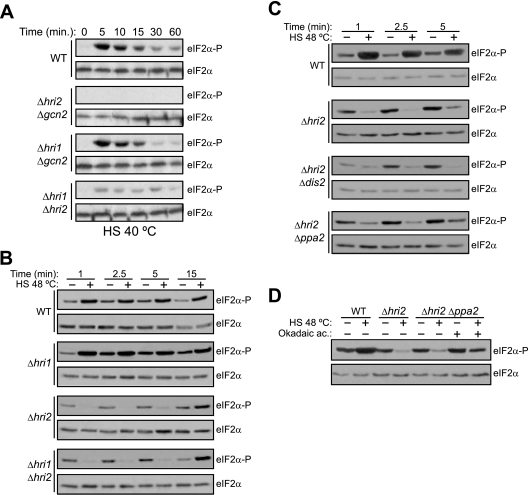
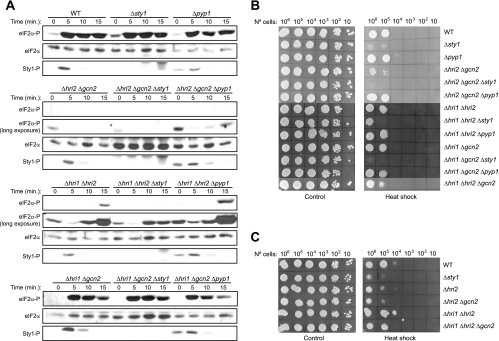
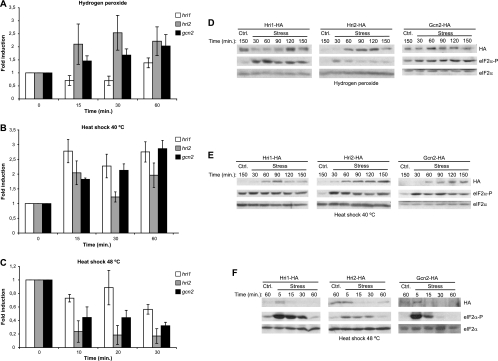
The mitogen-activated protein kinase (MAPK) Sty1 is essential for the regulation of transcriptional responses that promote cell survival in response to different types of environmental stimuli in Schizosaccharomyces pombe. In fission yeast, three distinct eukaryotic initiation factor 2alpha (eIF2alpha) kinases, two mammalian HRI-related protein kinases (Hri1 and Hri2) and the Gcn2 ortholog, regulate protein synthesis in response to cellular stress conditions. In this study, we demonstrate that both Hri1 and Hri2 exhibited an autokinase activity, specifically phosphorylated eIF2alpha, and functionally replaced the endogenous Saccharomyces cerevisiae Gcn2. We further show that Gcn2, but not Hri1 or Hri2, is activated early after exposure to hydrogen peroxide and methyl methanesulfonate (MMS). Cells lacking Gcn2 exhibit a later activation of Hri2. The activated MAPK Sty1 negatively regulates Gcn2 and Hri2 activities under oxidative stress but not in response to MMS. In contrast, Hri2 is the primary activated eIF2alpha kinase in response to heat shock. In this case, the activation of Sty1 appears to be transitory and does not contribute to the modulation of the eIF2alpha kinase stress pathway. In strains lacking Hri2, a type 2A protein phosphatase is activated soon after heat shock to reduce eIF2alpha phosphorylation. Finally, the MAPK Sty1, but not the eIF2alpha kinases, is essential for survival upon oxidative stress or heat shock, but not upon MMS treatment. These findings point to a regulatory coordination between the Sty1 MAPK and eIF2alpha kinase pathways for a particular range of stress responses.
Figures
Similar articles
-
New roles of the fission yeast eIF2α kinases Hri1 and Gcn2 in response to nutritional stress.J Cell Sci. 2013 Jul 15;126(Pt 14):3010-20. doi: 10.1242/jcs.118067. Epub 2013 May 17. J Cell Sci. 2013. PMID: 23687372
-
Phosphorylation of eukaryotic initiation factor 2 by heme-regulated inhibitor kinase-related protein kinases in Schizosaccharomyces pombe is important for fesistance to environmental stresses.Mol Cell Biol. 2002 Oct;22(20):7134-46. doi: 10.1128/MCB.22.20.7134-7146.2002. Mol Cell Biol. 2002. PMID: 12242291 Free PMC article.
-
Stress-activated protein kinase pathway functions to support protein synthesis and translational adaptation in response to environmental stress in fission yeast.Eukaryot Cell. 2005 Nov;4(11):1785-93. doi: 10.1128/EC.4.11.1785-1793.2005. Eukaryot Cell. 2005. PMID: 16278445 Free PMC article.
-
Rad3 and Sty1 function in Schizosaccharomyces pombe: an integrated response to DNA damage and environmental stress?Mol Microbiol. 2008 Apr;68(2):246-54. doi: 10.1111/j.1365-2958.2008.06147.x. Mol Microbiol. 2008. PMID: 18366437 Review.
-
Oxidative stress response pathways: Fission yeast as archetype.Crit Rev Microbiol. 2015;41(4):520-35. doi: 10.3109/1040841X.2013.870968. Crit Rev Microbiol. 2015. PMID: 24576188 Review.
Cited by
-
General amino acid control in fission yeast is regulated by a nonconserved transcription factor, with functions analogous to Gcn4/Atf4.Proc Natl Acad Sci U S A. 2018 Feb 20;115(8):E1829-E1838. doi: 10.1073/pnas.1713991115. Epub 2018 Feb 5. Proc Natl Acad Sci U S A. 2018. PMID: 29432178 Free PMC article.
-
Ribosome profiling reveals ribosome stalling on tryptophan codons and ribosome queuing upon oxidative stress in fission yeast.Nucleic Acids Res. 2021 Jan 11;49(1):383-399. doi: 10.1093/nar/gkaa1180. Nucleic Acids Res. 2021. PMID: 33313903 Free PMC article.
-
Cross Talk between eIF2α and eEF2 Phosphorylation Pathways Optimizes Translational Arrest in Response to Oxidative Stress.iScience. 2019 Oct 25;20:466-480. doi: 10.1016/j.isci.2019.09.031. Epub 2019 Sep 26. iScience. 2019. PMID: 31627132 Free PMC article.
-
Folding Mitochondrial-Mediated Cytosolic Proteostasis Into the Mitochondrial Unfolded Protein Response.Front Cell Dev Biol. 2021 Sep 23;9:715923. doi: 10.3389/fcell.2021.715923. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34631705 Free PMC article. Review.
-
The Fission Yeast Cell Integrity Pathway: A Functional Hub for Cell Survival upon Stress and Beyond.J Fungi (Basel). 2021 Dec 30;8(1):32. doi: 10.3390/jof8010032. J Fungi (Basel). 2021. PMID: 35049972 Free PMC article. Review.
References
-
- Bahler, J., J. Q. Wu, M. S. Longtine, N. G. Shah, A. McKenzie III, A. B. Steever, A. Wach, P. Philippsen, and J. R. Pringle. 1998. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast 14:943-951. - PubMed
-
- Berlanga, J. J., S. Herrero, and C. de Haro. 1998. Characterization of the hemin-sensitive eukaryotic initiation factor 2α kinase from mouse nonerythroid cells. J. Biol. Chem. 273:32340-32346. - PubMed
-
- Berlanga, J. J., J. Santoyo, and C. De Haro. 1999. Characterization of a mammalian homolog of the GCN2 eukaryotic initiation factor 2α kinase. Eur. J. Biochem. 265:754-762. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
