

Large structural change in isolated synaptic vesicles upon loading with neurotransmitter
- PMID: 19883601
- PMCID: PMC2770603
- DOI: 10.1016/j.bpj.2009.08.032
Large structural change in isolated synaptic vesicles upon loading with neurotransmitter
Abstract
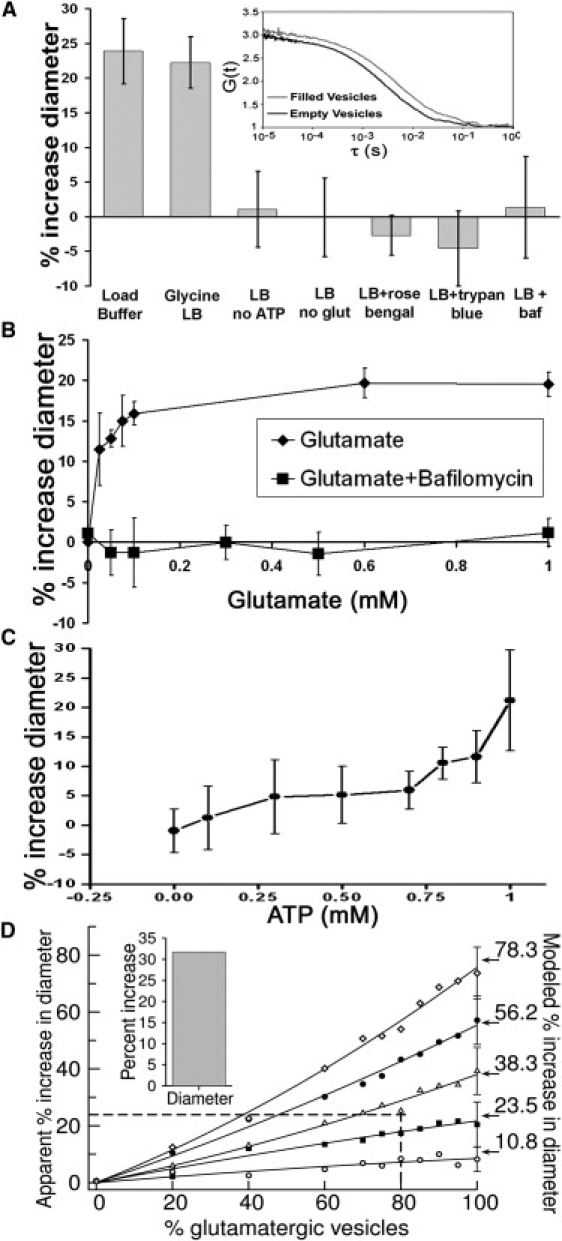
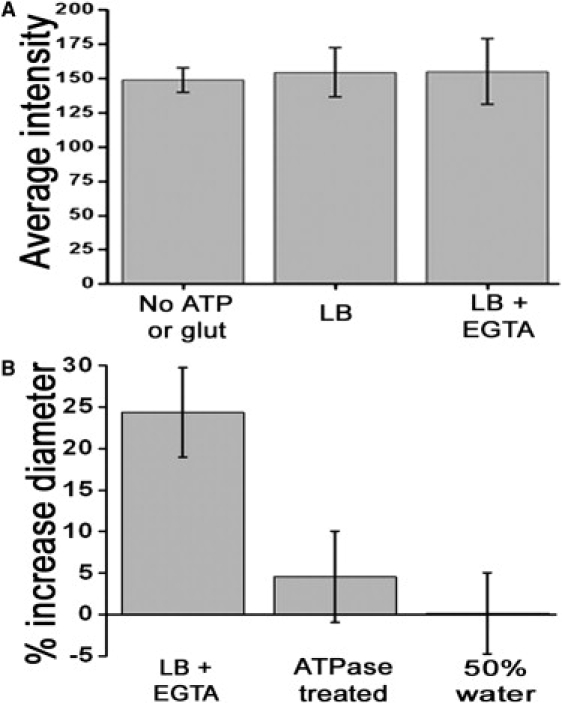
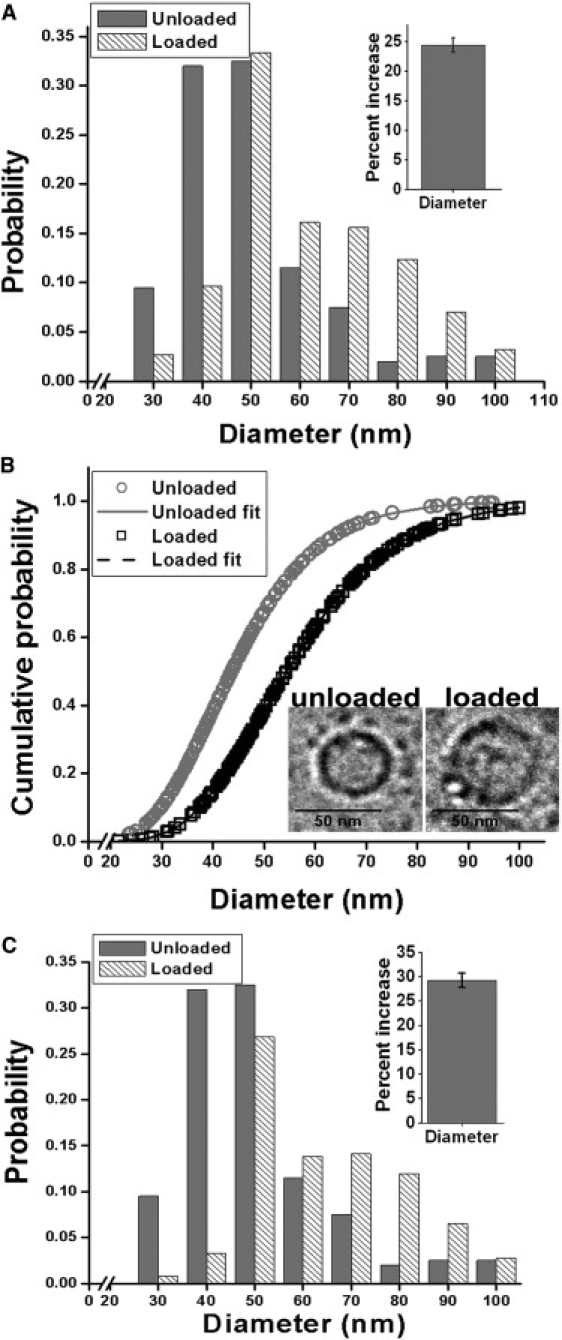
The size of a synaptic vesicle (SV) is generally thought to be determined by the amount of lipid and membrane protein it contains. Once formed, it is thought to remain constant in size. Using fluorescence correlation spectroscopy and cryogenic electron microscopy, we show that glutamatergic vesicles reversibly increase their size upon filling with glutamate. The increase ( approximately 25% in diameter) corresponds to an increase in surface area of approximately 50% and in volume of approximately 100%. This large size increase implies a large structural change in the SV upon loading with neurotransmitters. Vesicles lacking SV protein 2A (SV2A) did not manifest a change in size after loading with glutamate, indicating that SV2A is required for this phenomenon.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
