

Human serum albumin inhibits Abeta fibrillization through a "monomer-competitor" mechanism
- PMID: 19883602
- PMCID: PMC2770600
- DOI: 10.1016/j.bpj.2009.08.028
Human serum albumin inhibits Abeta fibrillization through a "monomer-competitor" mechanism
Abstract
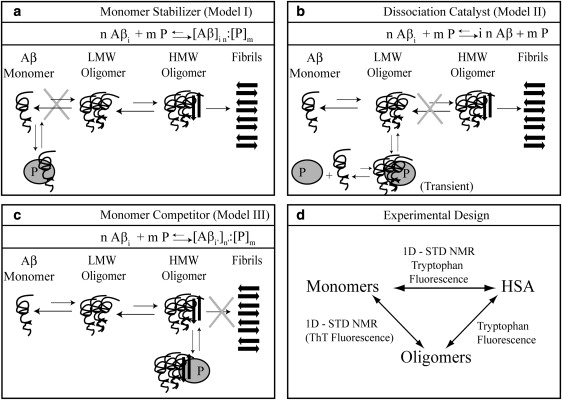
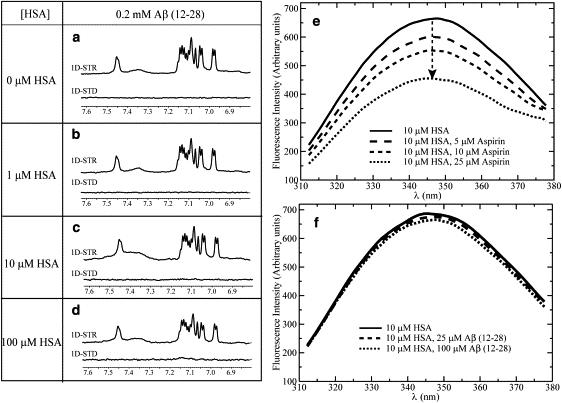
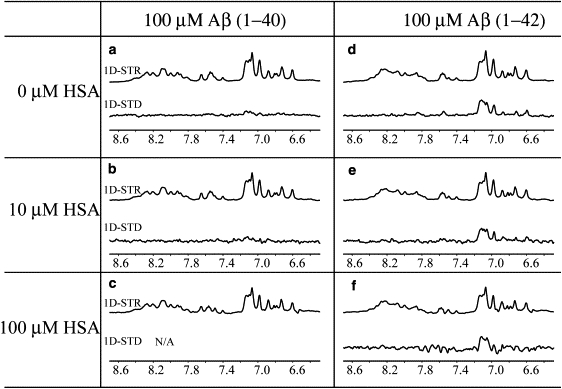
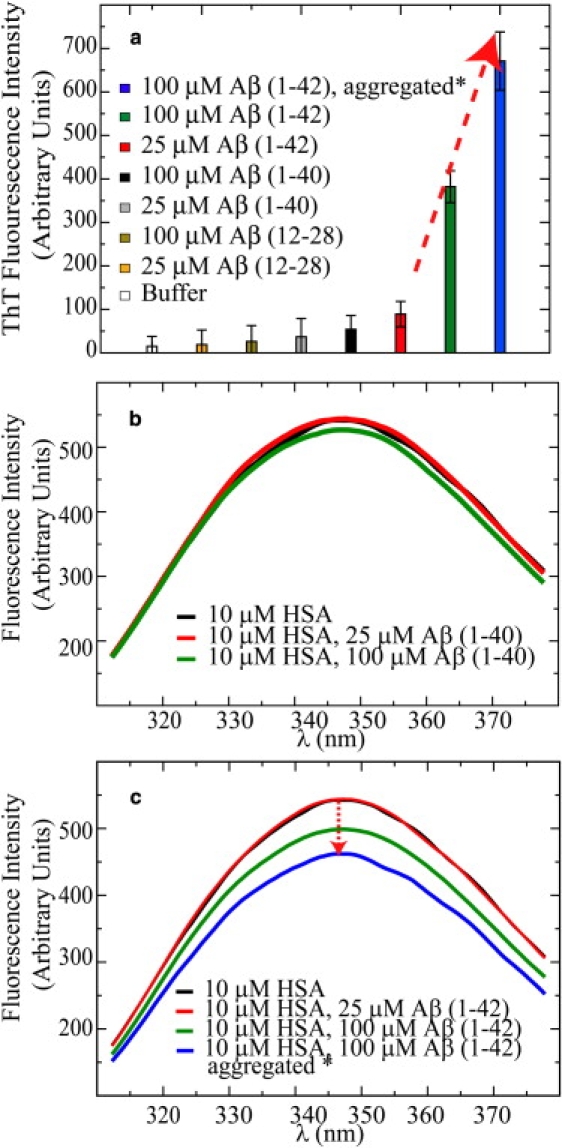
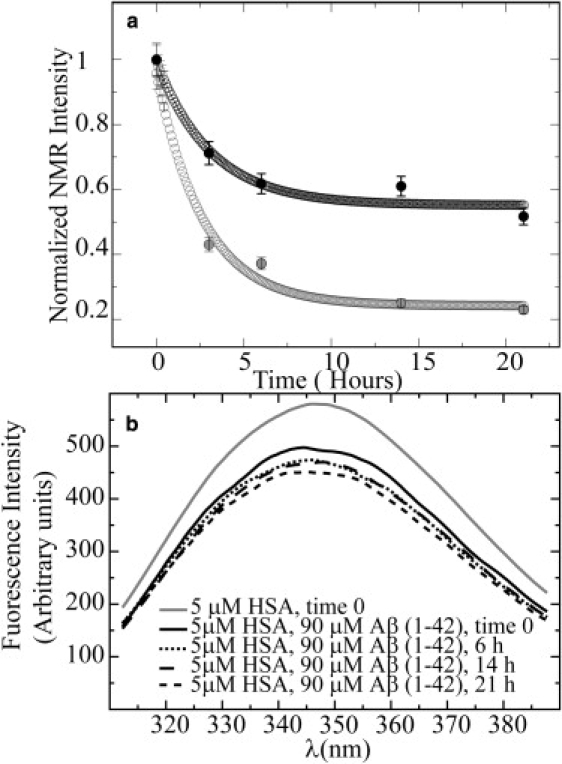
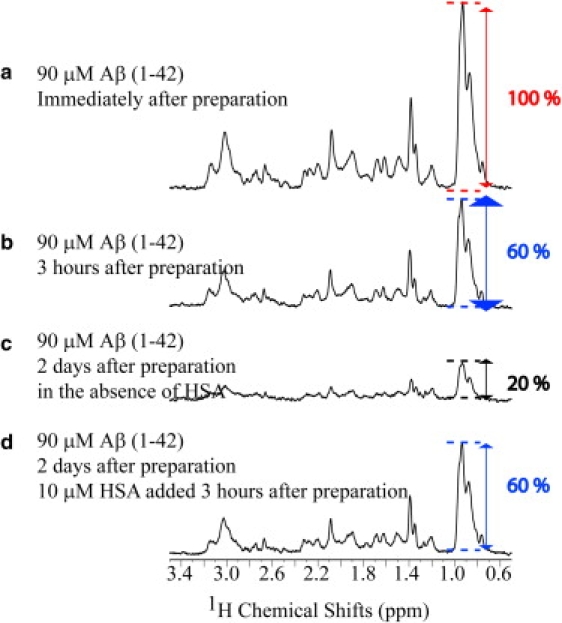
Human serum albumin (HSA) is not only a fatty acid and drug carrier protein, it is also a potent inhibitor of Abeta self-association in plasma. However, the mechanism underlying the inhibition of Abeta fibrillization by HSA is still not fully understood. We therefore investigated the Abeta-HSA system using a combined experimental strategy based on saturation transfer difference (STD) NMR and intrinsic albumin fluorescence experiments on three Abeta peptides with different aggregation propensities (i.e., Abeta(12-28), Abeta(1-40), and Abeta(1-42)). Our data consistently show that albumin selectively binds to cross-beta-structured Abeta oligomers as opposed to Abeta monomers. The HSA/Abeta oligomer complexes have K(D) values in the micromolar to submicromolar range and compete with the further addition of Abeta monomers to the Abeta assemblies, thus inhibiting fibril growth ("monomer competitor" model). Other putative mechanisms, according to which albumin acts as a "monomer stabilizer" or a "dissociation catalyst", are not supported by our data, thus resolving previous discrepancies in the literature regarding Abeta-HSA interactions. In addition, the model and the experimental approaches proposed here are anticipated to have broad relevance for the characterization of other systems that involve amyloidogenic peptides and oligomerization inhibitors.
Figures
References
-
- Cummings J.L. Alzheimer's disease. N. Engl. J. Med. 2004;351:56–67. - PubMed
-
- Haass C., Selkoe D.J. Cellular processing of β- amyloid precursor protein and the genesis of amyloid β -peptide. Cell. 1993;73:1039–1042. - PubMed
-
- Walsh D.M., Klyubin I., Fadeeva J.V., Cullen W.K., Anwyl R. Naturally secreted oligomers of the Alzheimer amyloid β-protein potently inhibit hippocampal long-term potentiation in vivo. Nature. 2002;416:535–539. - PubMed
-
- Dahlgren K.N., Manelli A.M., Stine W.K., Baker L.K., Jr., Krafft G.A. Oligomeric and fibrillar species of amyloid-β peptides differentially affect neuronal viability. J. Biol. Chem. 2002;277:32046–32053. - PubMed
-
- Zlokovic B.V. Clearing amyloid through the blood–brain barrier. J. Neurochem. 2004;89:807–811. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
