

Cyclodextrin overcomes deficient lysosome-to-endoplasmic reticulum transport of cholesterol in Niemann-Pick type C cells
- PMID: 19884502
- PMCID: PMC2780767
- DOI: 10.1073/pnas.0910916106
Cyclodextrin overcomes deficient lysosome-to-endoplasmic reticulum transport of cholesterol in Niemann-Pick type C cells
Abstract
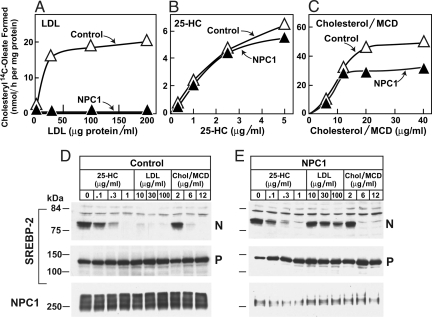
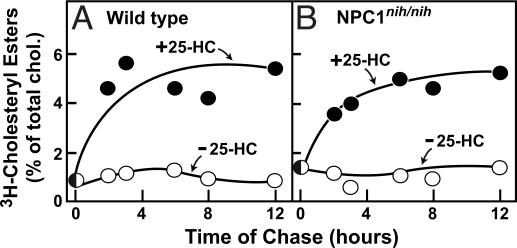
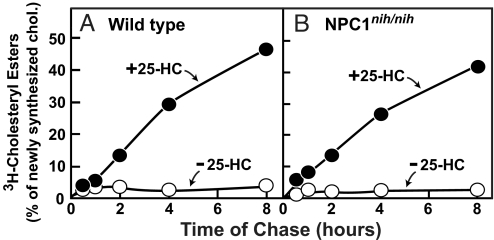
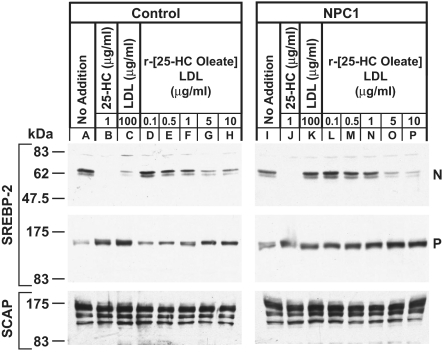
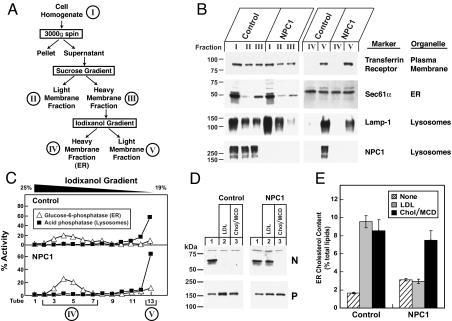
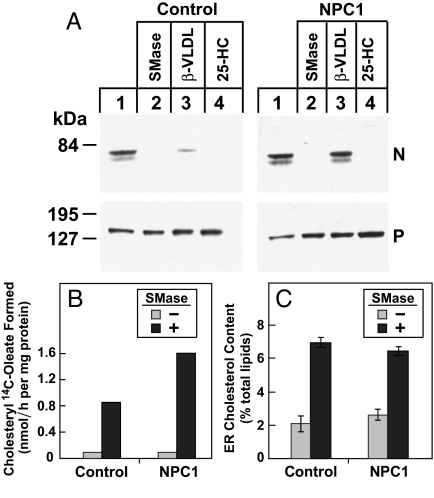
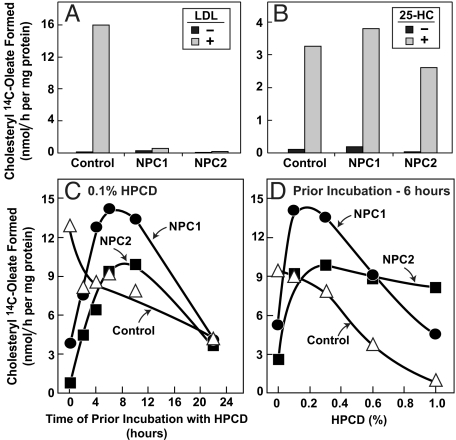
A handoff model has been proposed to explain the egress from lysosomes of cholesterol derived from receptor-mediated endocytosis of LDL. Cholesterol is first bound by soluble Niemann-Pick C2 (NPC2) protein, which hands off the cholesterol to the N-terminal domain of membrane-bound NPC1. Cells lacking NPC1 or NPC2 accumulate LDL-derived cholesterol in lysosomes and fail to deliver LDL cholesterol to the endoplasmic reticulum (ER) for esterification by acyl-CoA acyltransferase (ACAT) and for inhibition of sterol regulatory element-binding protein cleavage. Here, we support this model by showing that the cholesterol transport defect in NPC1 mutant cells is restricted to lysosomal export. Other cholesterol transport pathways appear normal, including the movement of cholesterol from the plasma membrane to the ER after treatment of cells with 25-hydroxycholesterol or sphingomyelinase. The NPC1 or NPC2 block in cholesterol delivery to the ER can be overcome by 2-hydroxypropyl-beta-cyclodextrin, which leads to a marked increase in ACAT-mediated cholesterol esterification. The buildup of cholesteryl esters in the cytosol is expected to be much less toxic than the buildup of free cholesterol in the lysosomes of patients with mutations in NPC1 or NPC2.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Pentchev PG, Vanier MT, Suzuki K, Patterson MC. In: The Metabolic and Molecular Bases of Inherited Disease. 7th Ed. Scriver CR, Beaudet AL, Sly WS, Valle D, editors. Vol II. New York: McGraw-Hill; 1995. pp. 2625–2639.
-
- Liscum L, Sturley SL. Intracellular trafficking of Niemann-Pick C proteins 1 and 2: Obligate components of subcellular lipid transport. Biochim Biophys Acta. 2004;1685:22–27. - PubMed
-
- Brown MS, Goldstein JL. A receptor-mediated pathway for cholesterol homeostasis. Science. 1986;232:34–47. - PubMed
-
- Liou H-L, et al. NPC2, the protein deficient in Niemann-Pick C2 disease, consists of multiple glycoforms that bind a variety of sterols. J Biol Chem. 2006;281:36710–36723. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
