

Calorie restriction increases fatty acid synthesis and whole body fat oxidation rates
- PMID: 19887594
- PMCID: PMC4056782
- DOI: 10.1152/ajpendo.00524.2009
Calorie restriction increases fatty acid synthesis and whole body fat oxidation rates
Abstract
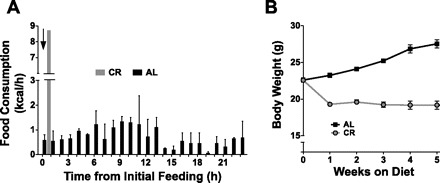
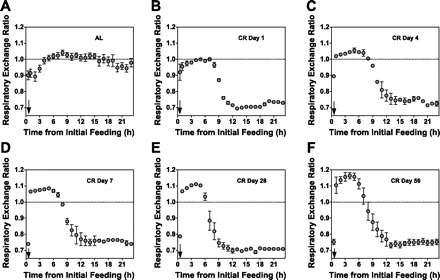
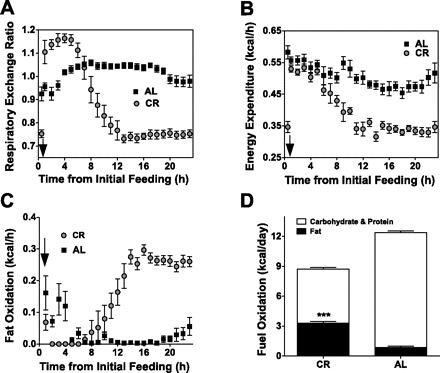
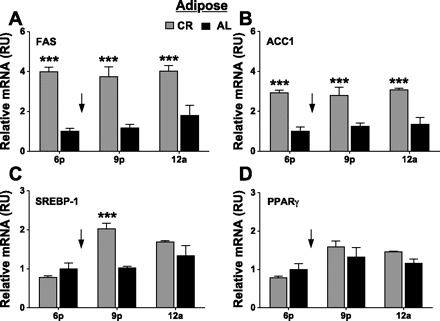
Calorie restriction (CR) increases longevity and retards the development of many chronic diseases, but the underlying metabolic signals are poorly understood. Increased fatty acid (FA) oxidation and reduced FA synthesis have been hypothesized to be important metabolic adaptations to CR. However, at metabolic steady state, FA oxidation must match FA intake plus synthesis; moreover, FA intake is low, not high, during CR. Therefore, it is not clear how FA dynamics are altered during CR. Accordingly, we measured food intake patterns, whole body fuel selection, endogenous FA synthesis, and gene expression in mice on CR. Within 2 days of CR being started, a shift to a cyclic, diurnal pattern of whole body FA metabolism occurred, with an initial phase of elevated endogenous FA synthesis [respiratory exchange ratio (RER) >1.10, lasting 4-6 h after food provision], followed by a prolonged phase of FA oxidation (RER = 0.70, lasting 18-20 h). CR mice oxidized four times as much fat per day as ad libitum (AL)-fed controls (367 +/- 19 vs. 97 +/- 14 mg/day, P < 0.001) despite reduced energy intake from fat. This increase in FA oxidation was balanced by a threefold increase in adipose tissue FA synthesis compared with AL. Expression of FA synthase and acetyl-CoA carboxylase mRNA were increased in adipose and liver in a time-dependent manner. We conclude that CR induces a surprising metabolic pattern characterized by periods of elevated FA synthesis alternating with periods of FA oxidation disproportionate to dietary FA intake. This pattern may have implications for oxidative damage and disease risk.
Figures



References
-
- Bassilian S, Ahmed S, Lim SK, Boros LG, Mao CS, Lee WN. Loss of regulation of lipogenesis in the Zucker diabetic rat. II. Changes in stearate and oleate synthesis. Am J Physiol Endocrinol Metab 282: E507–E513, 2002 - PubMed
-
- Berrigan D, Lavigne JA, Perkins SN, Nagy TR, Barrett JC, Hursting SD. Phenotypic effects of calorie restriction and insulin-like growth factor-1 treatment on body composition and bone mineral density of C57BL/6 mice: implications for cancer prevention. In Vivo 19: 667–674, 2005 - PubMed
-
- Brochmann EJ, Duarte ME, Zaidi HA, Murray SS. Effects of dietary restriction on total body, femoral, and vertebral bone in SENCAR, C57BL/6, and DBA/2 mice. Metabolism 52: 1265–1273, 2003 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources