

Characterization of Lassa virus glycoprotein oligomerization and influence of cholesterol on virus replication
- PMID: 19889753
- PMCID: PMC2798336
- DOI: 10.1128/JVI.02039-09
Characterization of Lassa virus glycoprotein oligomerization and influence of cholesterol on virus replication
Abstract
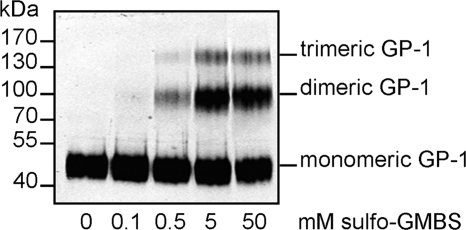
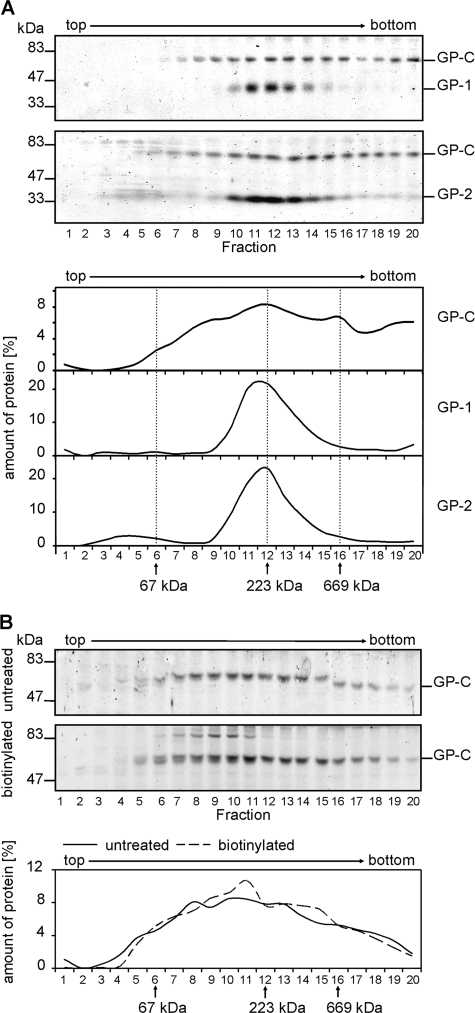
Mature glycoprotein spikes are inserted in the Lassa virus envelope and consist of the distal subunit GP-1, the transmembrane-spanning subunit GP-2, and the signal peptide, which originate from the precursor glycoprotein pre-GP-C by proteolytic processing. In this study, we analyzed the oligomeric structure of the viral surface glycoprotein. Chemical cross-linking studies of mature glycoprotein spikes from purified virus revealed the formation of trimers. Interestingly, sucrose density gradient analysis of cellularly expressed glycoprotein showed that in contrast to trimeric mature glycoprotein complexes, the noncleaved glycoprotein forms monomers and oligomers spanning a wide size range, indicating that maturation cleavage of GP by the cellular subtilase SKI-1/S1P is critical for formation of the correct oligomeric state. To shed light on a potential relation between cholesterol and GP trimer stability, we performed cholesterol depletion experiments. Although depletion of cholesterol had no effect on trimerization of the glycoprotein spike complex, our studies revealed that the cholesterol content of the viral envelope is important for the infectivity of Lassa virus. Analyses of the distribution of viral proteins in cholesterol-rich detergent-resistant membrane areas showed that Lassa virus buds from membrane areas other than those responsible for impaired infectivity due to cholesterol depletion of lipid rafts. Thus, derivation of the viral envelope from cholesterol-rich membrane areas is not a prerequisite for the impact of cholesterol on virus infectivity.
Figures
References
-
- Aizaki, H., K. Morikawa, M. Fukasawa, H. Hara, Y. Inoue, H. Tani, K. Saito, M. Nishijima, K. Hanada, Y. Matsuura, M. M. Lai, T. Miyamura, T. Wakita, and T. Suzuki. 2008. Critical role of virion-associated cholesterol and sphingolipid in hepatitis C virus infection. J. Virol. 82:5715-5724. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
