

Small G proteins in islet beta-cell function
- PMID: 19890090
- PMCID: PMC2852207
- DOI: 10.1210/er.2009-0022
Small G proteins in islet beta-cell function
Abstract
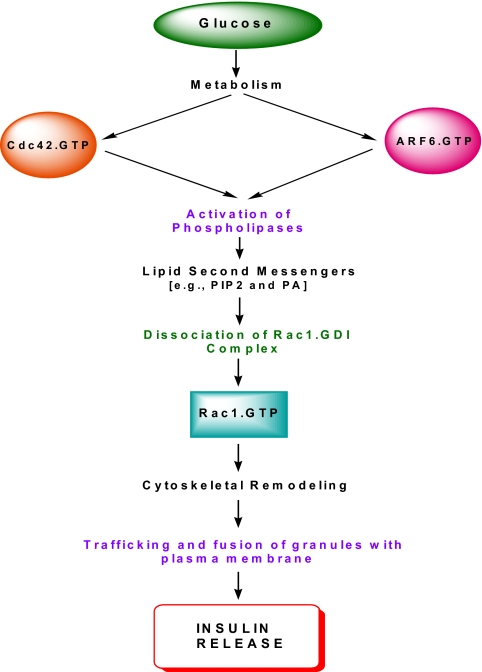
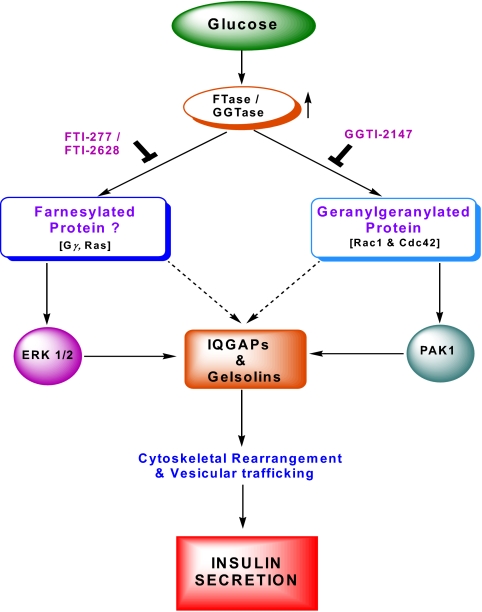
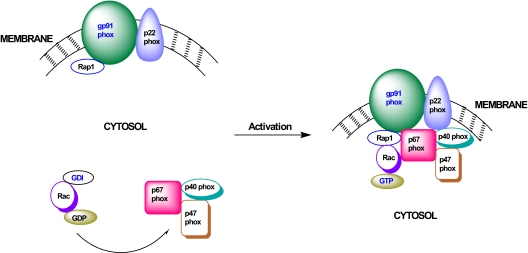
Glucose-stimulated insulin secretion from the islet beta-cell involves a sequence of metabolic events and an interplay between a wide range of signaling pathways leading to the generation of second messengers (e.g., cyclic nucleotides, adenine and guanine nucleotides, soluble lipid messengers) and mobilization of calcium ions. Consequent to the generation of necessary signals, the insulin-laden secretory granules are transported from distal sites to the plasma membrane for fusion and release of their cargo into the circulation. The secretory granule transport underlies precise changes in cytoskeletal architecture involving a well-coordinated cross-talk between various signaling proteins, including small molecular mass GTP-binding proteins (G proteins) and their respective effector proteins. The purpose of this article is to provide an overview of current understanding of the identity of small G proteins (e.g., Cdc42, Rac1, and ARF-6) and their corresponding regulatory factors (e.g., GDP/GTP-exchange factors, GDP-dissociation inhibitors) in the pancreatic beta-cell. Plausible mechanisms underlying regulation of these signaling proteins by insulin secretagogues are also discussed. In addition to their positive modulatory roles, certain small G proteins also contribute to the metabolic dysfunction and demise of the islet beta-cell seen in in vitro and in vivo models of impaired insulin secretion and diabetes. Emerging evidence also suggests significant insulin secretory abnormalities in small G protein knockout animals, further emphasizing vital roles for these proteins in normal health and function of the islet beta-cell. Potential significance of these experimental observations from multiple laboratories and possible avenues for future research in this area of islet research are highlighted.
Figures
Similar articles
-
Role of G-proteins in islet function in health and diabetes.Diabetes Obes Metab. 2017 Sep;19 Suppl 1(Suppl 1):63-75. doi: 10.1111/dom.13011. Diabetes Obes Metab. 2017. PMID: 28880478 Free PMC article. Review.
-
Arf nucleotide binding site opener [ARNO] promotes sequential activation of Arf6, Cdc42 and Rac1 and insulin secretion in INS 832/13 β-cells and rat islets.Biochem Pharmacol. 2011 Apr 15;81(8):1016-27. doi: 10.1016/j.bcp.2011.01.006. Epub 2011 Jan 26. Biochem Pharmacol. 2011. PMID: 21276423 Free PMC article.
-
Emerging Roles of Small GTPases in Islet β-Cell Function.Cells. 2021 Jun 15;10(6):1503. doi: 10.3390/cells10061503. Cells. 2021. PMID: 34203728 Free PMC article. Review.
-
Friendly, and not so friendly, roles of Rac1 in islet β-cell function: lessons learnt from pharmacological and molecular biological approaches.Biochem Pharmacol. 2011 Apr 15;81(8):965-75. doi: 10.1016/j.bcp.2011.01.013. Epub 2011 Feb 12. Biochem Pharmacol. 2011. PMID: 21300027 Free PMC article.
-
Tiam1/Vav2-Rac1 axis: A tug-of-war between islet function and dysfunction.Biochem Pharmacol. 2017 May 15;132:9-17. doi: 10.1016/j.bcp.2017.02.007. Epub 2017 Feb 13. Biochem Pharmacol. 2017. PMID: 28202288 Free PMC article. Review.
Cited by
-
Rab-geranylgeranyl transferase regulates glucose-stimulated insulin secretion from pancreatic β cells.Islets. 2012 Sep-Oct;4(5):354-8. doi: 10.4161/isl.22538. Epub 2012 Sep 1. Islets. 2012. PMID: 23114750 Free PMC article.
-
Statin Short-term Inhibition of Insulin Sensitivity and Secretion During Acute Phase of ST-Elevation Myocardial Infarction.Sci Rep. 2019 Nov 8;9(1):16401. doi: 10.1038/s41598-019-52111-x. Sci Rep. 2019. PMID: 31704948 Free PMC article. Clinical Trial.
-
Connecting pancreatic islet lipid metabolism with insulin secretion and the development of type 2 diabetes.Ann N Y Acad Sci. 2020 Feb;1461(1):53-72. doi: 10.1111/nyas.14037. Epub 2019 Apr 2. Ann N Y Acad Sci. 2020. PMID: 30937918 Free PMC article. Review.
-
SAD-A Promotes Glucose-Stimulated Insulin Secretion Through Phosphorylation and Inhibition of GDIα in Male Islet β Cells.Endocrinology. 2018 Aug 1;159(8):3036-3047. doi: 10.1210/en.2017-03243. Endocrinology. 2018. PMID: 29873699 Free PMC article.
-
On a sugary-relationship between caspases and lamins.Cell Cycle. 2014;13(24):3787-8. doi: 10.4161/15384101.2014.989948. Cell Cycle. 2014. PMID: 25470565 Free PMC article. No abstract available.
References
-
- Malaisse WJ 1972 Hormonal and environmental modification of islet activity. In: Steiner DF, Frankel N, eds. Handbook of physiology. New York: American Physiological Society; 237–260
-
- Prentki M, Matschinsky FM 1987 Calcium, cAMP, and phospholipid-derived messengers in coupling mechanisms of insulin secretion. Physiol Rev 67:1185–1248 - PubMed
-
- MacDonald MJ 1990 Elusive proximal signals of β-cells for insulin secretion. Diabetes 39:1461–1466 - PubMed
-
- Laychock SG 1990 Glucose metabolism, second messengers and insulin secretion. Life Sci 47:2307–2316 - PubMed
-
- Newgard CB, McGarry JD 1995 Metabolic coupling factors in pancreatic β-cell signal transduction. Annu Rev Biochem 64:689–719 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
