

Local maternal control of seed size by KLUH/CYP78A5-dependent growth signaling
- PMID: 19892740
- PMCID: PMC2785301
- DOI: 10.1073/pnas.0907024106
Local maternal control of seed size by KLUH/CYP78A5-dependent growth signaling
Abstract
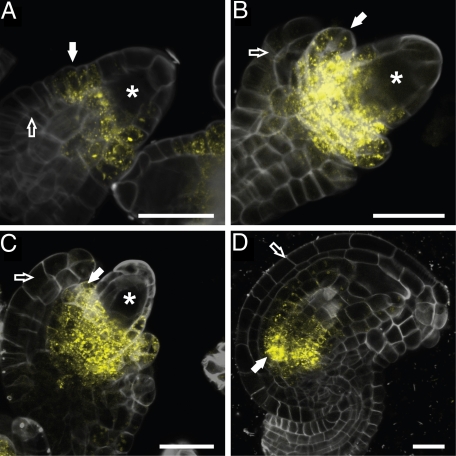
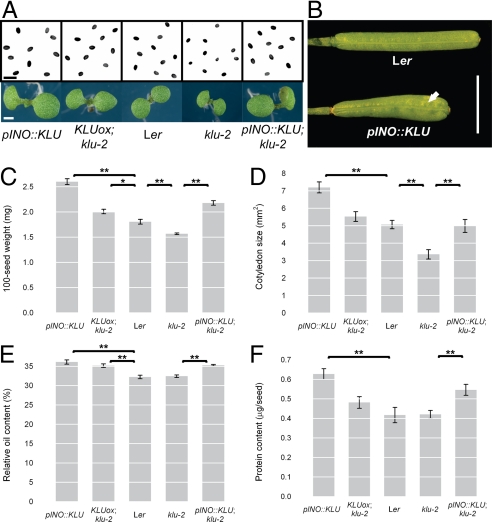
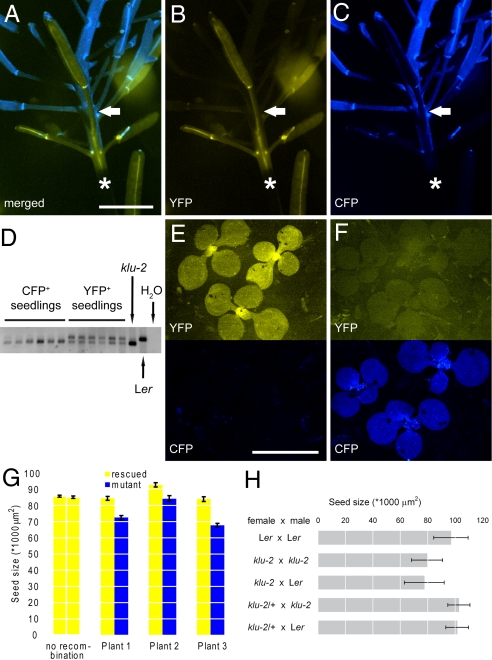
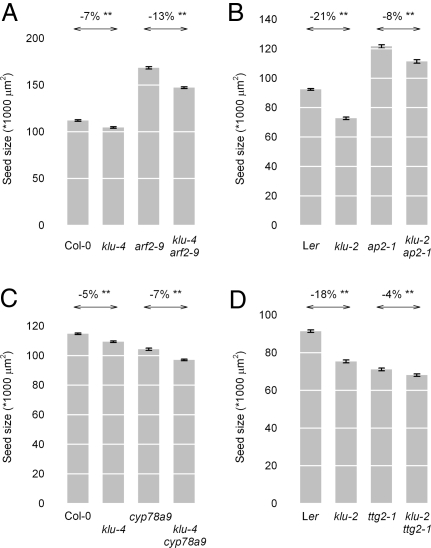
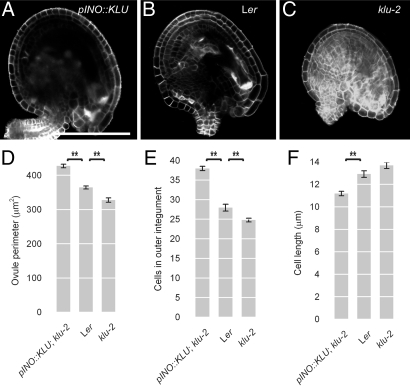
Seed development in plants involves the coordinated growth of the embryo, endosperm, and maternal tissue. Several genes have been identified that influence seed size by acting maternally, such as AUXIN RESPONSE FACTOR2, APETALA2, and DA1. However, given the lack of gain-of-function effects of these genes on seed size, it is unclear whether their activity levels are limiting in WT plants and whether they could thus be used to regulate seed size in development or evolution. Also, whether the altered seed sizes reflect local gene activity or global physiological changes is unknown. Here, we demonstrate that the cytochrome P450 KLUH (KLU) regulates seed size. KLU acts locally in developing flowers to promote seed growth, and its activity level is limiting for seed growth in WT. KLU is expressed in the inner integument of developing ovules, where it non-cell autonomously stimulates cell proliferation, thus determining the growth potential of the seed coat and seed. A KLU-induced increase in seed size leads to larger seedlings and higher relative oil content of the seeds. Genetic analyses indicate that KLU acts independently of other tested maternal factors that influence integument cell proliferation. Thus, the level of KLU-dependent growth factor signaling determines size in ovules and seeds, suggesting this pathway as a target for crop improvement.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Harper JL, Lovell PH, Moore KG. The shapes and sizes of seeds. Annu Rev Ecol Syst. 1970;1:327–356.
-
- Venable DL. Size-number trade-offs and the variation of seed size with plant resource status. Am Nat. 1992;140:287–304.
-
- Shomura A, et al. Deletion in a gene associated with grain size increased yields during rice domestication. Nat Genet. 2008;40:1023–1028. - PubMed
-
- Berger F, Grini PE, Schnittger A. Endosperm: An integrator of seed growth and development. Curr Opin Plant Biol. 2006;9:664–670. - PubMed
-
- Colombo L, Battaglia R, Kater MM. Arabidopsis ovule development and its evolutionary conservation. Trends Plants Sci. 2008;13:444–450. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
